熱線:021-66110810,66110819
手機:13564362870
熱線:021-66110810,66110819
手機:13564362870
3.5循環中硫化氫水平的降低導致缺氧性腦損傷小鼠紅細胞中2,3-BPG和P50水平的增加
降低循環硫化氫水平有助于增加缺氧性腦損傷小鼠紅細胞中的2,3-BPG和P50水平。與先前的缺氧結果一致。研究顯示,小鼠在缺氧條件下,紅細胞2,3-BPG水平在一天內升高,并在缺氧持續3天期間保持高水平(圖8(a))。此外,P50在缺氧一天后增加約15%,并在缺氧第三天繼續上升(圖8(b))。我們還檢測了小鼠在缺氧刺激下循環系統中硫化氫的水平。如圖8(c)所示,缺氧期間循環系統中的硫化氫水平在第一天顯著降低,并在第三天進一步減少。紅細胞、肝臟和腎臟中的CSE表達水平在缺氧期間也顯著降低。有趣的是,GYY4137處理逆轉了缺氧引起的紅細胞2,3-BPG和P50水平升高(圖8(d)和8(e))。隨后,我們測試了缺氧對紅細胞膜Hb含量、胞質BPGM活性以及BPGM從膜到胞質轉運的影響,以及缺氧期間硫化氫干預的效果。如預期,與正常氧環境下的小鼠相比,缺氧暴露組小鼠的紅細胞中血紅素含量顯著升高(圖8(f))。與此一致的是,缺氧組小鼠紅細胞的胞質BPGM活性顯著增強(圖8(g)),而膜BPGM水平顯著降低,胞質BPGM水平則顯著升高(圖8(h))。在缺氧條件下,GYY4137處理可降低紅細胞膜血紅素含量,同時提升胞質BPGM活性和膜BPGM水平,降低胞質BPGM水平(圖8(f)–8(h))。此外,缺氧組小鼠的血紅蛋白硫水合水平降低,而GYY4137處理可逆轉這一現象(圖8(i))。
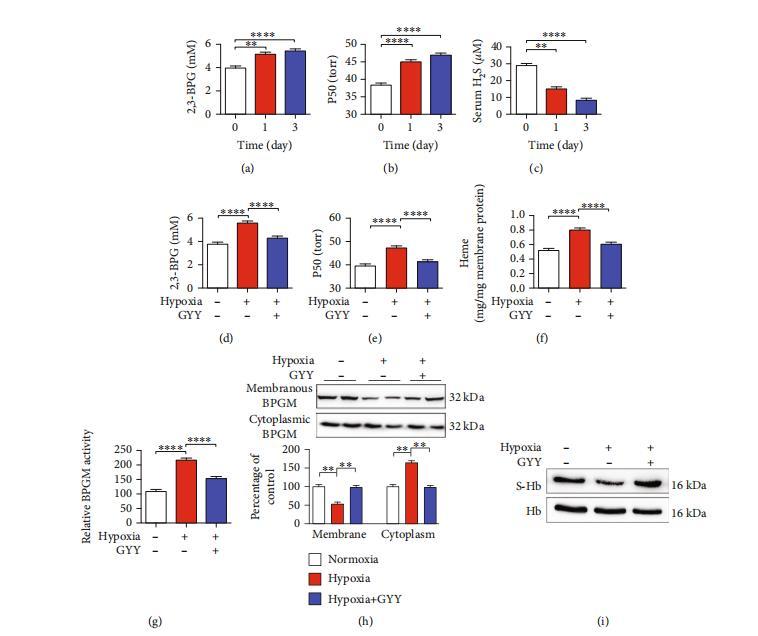
圖8:硫化氫產量的減少導致小鼠紅細胞中缺氧誘導的2,3-BPG和P50水平升高,以及BPGM和Hb在膜與細胞質之間的易位。(a–c)缺氧對2,3-BPG、P50和硫化氫水平的影響。野生型小鼠暴露于缺氧(10%氧氣)指定時間。采集血液樣本以測定紅細胞2,3-BPG(a)和P50(b)水平,以及血漿中的硫化氫水平(c)。數據以均值±標準誤表示(n=10)。??p<0.01,????p<0.0001。(d–h)GYY4137對缺氧誘導的2,3-BPG和P50水平升高以及BPGM和Hb從膜到細胞質易位的影響。野生型小鼠暴露于缺氧(10%氧氣)并給予GYY4137或生理鹽水。缺氧和GYY413處理24小時后,采集血液樣本以測定紅細胞2,3-BPG(d)、P50(e)、膜血紅素濃度(f)、細胞質BPGM活性(g)以及膜和細胞質BPGM水平(h)。數據以均值±標準誤表示(n=10)。????p<0.0001。(h)小鼠紅細胞中Hb的S-硫水合的代表性圖像。野生型小鼠暴露于缺氧(10%氧氣)并給予GYY4137(50 mg/kg)或生理鹽水。缺氧和GYY413處理24小時后,采集血液樣本以使用生物素開關技術測定Hb的S-硫水合。
討論
盡管硫化氫在眾多細胞和組織中發揮重要作用,但其在紅細胞中的功能仍不明確。在本研究中,我們從功能上揭示了硫化氫在控制2,3-BPG生成中起關鍵作用,并在常氧條件下維持血紅蛋白-氧合血紅蛋白(Hb-O2)親和力在正常范圍內。從機制上,我們發現內源性硫化氫抑制血紅蛋白向細胞膜的轉運,從而在常氧條件下維持BPGM主要錨定于細胞膜、2,3-BPG水平正常以及Hb-O2結合親和力。這些研究進一步使我們發現,外周組織中缺氧誘導的硫化氫生成減少導致小鼠紅細胞2,3-BPG水平升高和Hb-O2結合親和力降低。綜上所述,我們的發現揭示了內源性硫化氫在紅細胞中先前未被認識的作用,并闡明了硫化氫在常氧和缺氧條件下調節2,3-BPG和Hb-O2結合親和力的分子機制(圖9)。硫化氫一直被認為是重要的氣體信號分子,在健康和疾病中參與多種細胞和組織功能。
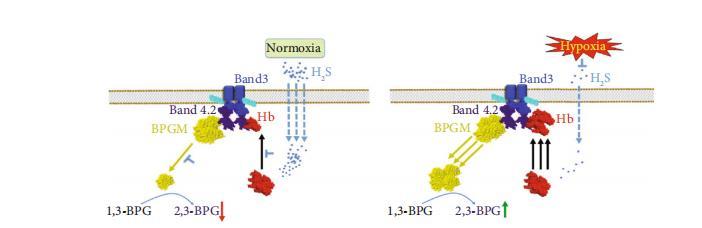
圖9:硫化氫在常氧和缺氧條件下調節2,3-BPG和Hb-O2結合親和力的潛在機制整合方案。硫化氫抑制Hb從細胞質向膜的轉運,進而促進BPGM錨定于膜上,從而防止常氧條件下2,3-BPG水平的升高。在缺氧期間,硫化氫水平的降低促進了Hb錨定于膜上,并促使BPGM從膜上釋放到細胞質中,最終導致紅細胞中2,3-BPG生成的增加。
關于硫化氫生成對缺氧刺激的響應變化可能存在爭議。一些研究表明,缺氧會促進某些組織(如胃腸道組織和頸動脈體)中硫化氫的生成,且這些組織中通常可檢測到PO2值和硫化氫濃度;因此,硫化氫被認為可以作為氧氣傳感器。相比之下,許多其他研究報道缺氧會抑制多種組織中硫化氫的生成,并導致循環系統中硫化氫水平下降。D‘Alessandro等人最近報告稱,高海拔會導致健康志愿者體內硫化氫水平立即下降。與此一致的是,我們驗證了硫化氫水平顯著在暴露于缺氧環境的小鼠中,我們發現其硫化氫水平降低,這與人類在高海拔環境下的情況相似。通過使用CSE基因修飾的小鼠模型,我們發現CSE產生的內源性硫化氫對2,3-BPG的生成具有持續抑制作用,并在常氧條件下維持Hb-O2結合親和力。盡管已有研究表明2,3-BPG和硫化氫在人類高海拔研究中會發生改變,但硫化氫是否在缺氧條件下調節2,3-BPG水平仍不明確。
與CSE缺陷小鼠類似,我們發現缺氧介導的硫化氫減少會導致2,3-BPG的誘導生成和Hb-O2結合親和力的降低。因此,我們的研究結果為紅細胞生理學和病理學提供了重要的新見解,揭示了硫化氫在常氧條件下維持2,3-BPG水平和Hb-O2結合親和力正常范圍的重要作用,以及硫化氫水平降低在促進2,3-BPG生成、從而降低Hb-O2結合親和力方面的關鍵作用,這可能是缺氧期間優化組織供氧的適應性機制。紅細胞2,3-BPG的形成受多種信號通路調控,其水平已知在缺氧條件下被誘導。它可被糖酵解酶的高活性刺激,從而增加葡萄糖被分流至BPGM反應。最近,Sun等人研究表明,鞘氨醇-1-磷酸(S1P)可增強GAPDH活性,從而促進紅細胞中2,3-BPG的生成。BPGM是2,3-BPG生成的關鍵限速酶。
最新研究顯示,在缺氧條件下,腺苷信號通路下游的硫氧還蛋白(AMPK)可激活BPGM。然而,硫化氫如何調控2,3-BPG生成仍不明確,直到我們證實了硫化氫在常氧和缺氧條件下對2,3-BPG生成及血紅蛋白-氧結合的重要性。值得注意的是,血紅蛋白與膜上帶3的胞質結構域結合,促使磷酸果糖激酶、醛縮酶和GAPDH等糖酵解酶從膜釋放到胞質,從而增強糖酵解作用,進而促進缺氧條件下2,3-BPG的生成。除血紅蛋白和糖酵解酶外,帶4.2、錨蛋白和光譜蛋白等多種蛋白質與紅細胞膜上的帶3相互作用,形成稱為帶3巨復合體的單一復合物。據我們所知,BPGM在紅細胞中是否介導膜與胞質間的轉運尚未見報道。通過多學科生化與遺傳學方法,我們揭示了由硫化氫介導的BPGM活性及2,3-BPG生成的調控機制。
具體而言,通過MS方法,我們從與BPGM相互作用的蛋白質中鑒定出4.2號帶蛋白、錨蛋白、光譜蛋白和3號帶蛋白,表明BPGM與紅細胞膜中的3號帶大復合體相關。利用紅細胞血影膜模型,我們揭示了BPGM在膜與細胞質之間的轉運過程,且BPGM與Hb的轉運相互關聯。此外,我們提供了堅實的遺傳學證據和藥理學證據,證明硫化氫是抑制Hb錨定到膜和BPGM從膜釋放的重要因素,從而維持2,3-BPG水平和Hb-O2結合親和力的正常范圍。紅細胞在適應低氧環境方面發揮著關鍵作用,這與其在氧氣運輸和遞送中的重要作用一致。
改善氧氣釋放是成熟紅細胞適應低氧的最重要適應之一。然而,對紅細胞在體內適應低氧的機制理解仍不完整。最近的研究表明,紅細胞鞘氨醇激酶1活性的升高介導的S1P增加是通過誘導氧氣釋放來實現低氧適應的。Liu等人顯示,5′AMP激活的蛋白激酶(AMPK)通過磷酸化和激活2,3-BPG變位酶介導低氧誘導的2,3-BPG生成。在本研究中,我們證明了硫化氫的下調導致低氧條件下Hb-O2結合親和力降低,從而揭示了紅細胞適應低氧的新型分子機制。
本研究的局限性在于,硫化氫調節紅細胞膜與細胞質之間Hb和BPGM轉運的機制仍有待進一步闡明。H2S誘導的翻譯后S-硫水合修飾已被證實對鉀通道和NF-κB等多種蛋白質的功能調控具有重要作用。最近,Zhao等人證明S-硫水合的Kelch樣ECH相關蛋白1(Keap1)和核因子紅系2相關因子(Nrf2)對硫化氫在應對多種損傷刺激時對肝組織的保護作用至關重要。
通過結合小鼠遺傳學研究與多種分子生物學操作,他們提供了Keap1的Cys151在硫化氫處理后發生硫水合的證據。盡管我們發現Hb的α鏈中的Cys104也發生了硫水合,但其是否對硫化氫調控Hb-O2親和力具有關鍵作用仍需進一步驗證。值得注意的是,Vitvitsky等人報道血紅蛋白可結合硫化氫并將其氧化為硫代硫酸鹽和水合多硫化物的混合物。該反應是否參與Hb-O2親和力的調控尚不明確。盡管如此,硫化氫調控紅細胞中Hb和BPGM轉運的機制仍需未來研究進一步闡明。
結論
總之,本研究揭示了以下新發現:硫化氫對于維持正常氧分壓下2,3-BPG水平和Hb-O2結合親和力的正常范圍至關重要,而硫化氫的下調是低氧條件下促進2,3-BPG生成的關鍵信號分子。鑒于所有活細胞都需要氧氣來適應生理和病理應激條件下的生存、增殖和分化,硫化氫介導的Hb-O2結合親和力變化很可能是對各種刺激的重要功能反應。
 相關新聞
相關新聞