熱線:021-66110810,66110819
手機:13564362870
熱線:021-66110810,66110819
手機:13564362870
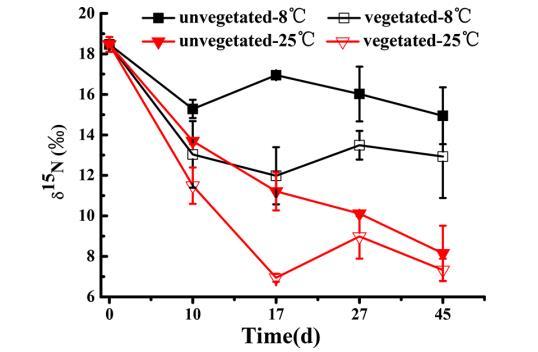
氮同化。 絮凝-覆蓋處理后,所有柱中沉積物樣品的δ15N富集度為18.47‰。該值在實驗過程中持續下降,在無植被-8°C、有植被-8°C、無植被-25°C和有植被-25°C組中分別達到約14.9‰、12.9‰、8.1‰和7.3‰。通常,在兩種培養溫度下,從第10天開始,有植被組沉積物中的δ15N顯著低于無植被組。此外,從第10天到第45天,有植被組和無植被組在25°C下的δ15N均顯著低于8°C下的值(P<0.05)。
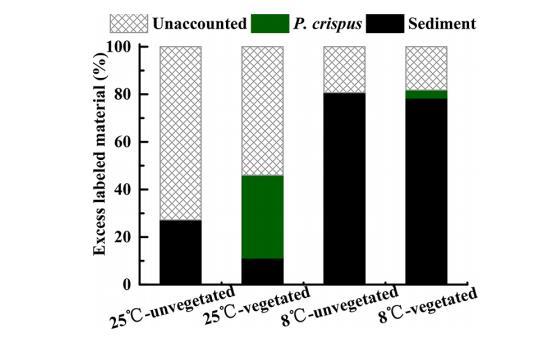
絮凝前,每個柱中填充了1.38 μmol 15N/g標記藻類。在實驗結束時對每個系統中過量15N的質量平衡進行了計算。在25°C下收集的蓖齒眼子菜中的過量15N(0.48 μmol 15N/g)比在8°C下(0.045 μmol 15N/g)高10倍。在8°C下培養的系統中,約有20%的過量15N未被計入。然而,在25°C下培養的柱中,約有55-75%的過量15N未被計入。
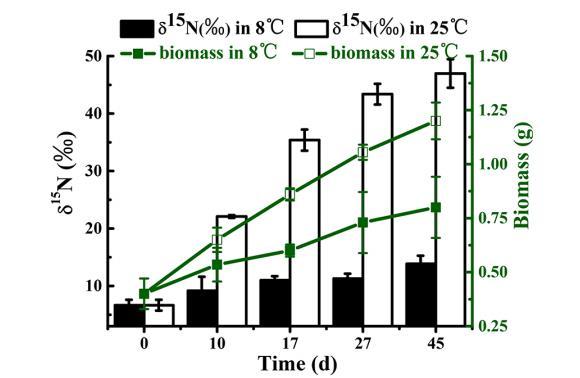
此外,在實驗期間,25°C下蓖齒眼子菜中δ15N的增加速率是8°C下的五倍。15N富集度的增加與蓖齒眼子菜生物量的增加顯著相關(r=0.915, P<0.05),其中在25°C下生長的蓖齒眼子菜生物量是在8°C下生長的兩倍。
討論
使用改性粘土/土壤沉降有害藻華。在沉水植物重新出現之前,去除和管理水華(尤其是藍藻水華)的生長是恢復富營養化湖泊的重要步驟。在本研究中,選擇改性土壤來加速藻華的沉降。土壤顆粒為藻生物量提供了足夠的壓載物,以抵消水柱中銅綠微囊藻細胞的浮力。藻絮體沉降到柱底部,而銅綠微囊藻主要懸浮在對照組水柱中。盡管在施用改性土壤后,F-no capping和F-capping處理組在每個溫度下的葉綠素a濃度顯示出相似的下降趨勢,但F-no capping組每個采樣點的葉綠素a濃度略高于F-capping組。這歸因于在F-no capping柱中存活的銅綠微囊藻細胞比在F-capping柱中更多。存活的藻生物量可能返回到水柱中,特別是在淺水區,那里會發生風和波浪引起的湍流。
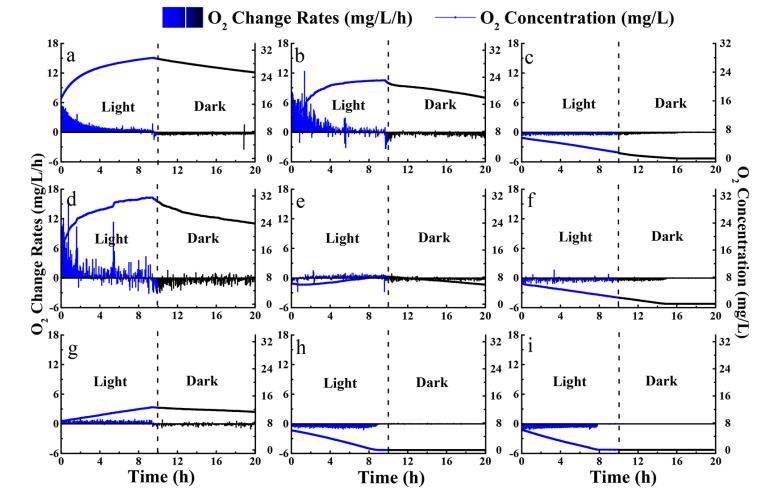
改性土壤絮凝和天然土壤覆蓋對銅綠微囊藻細胞造成的損害很小,這反映在第0天完整的細胞形態和正常的光合作用及呼吸作用上。這有助于目視觀察,在絮體周圍沒有出現均勻的綠色或黃色,這表明在此類實驗室實驗中發生了細胞裂解。然而,在進一步的研究中應進行其他分析,如溶解性葉綠素a、毒素和核酸,以證明目視觀察的結果。絮凝-覆蓋處理后觀察到的完整藻細胞對于防止細胞內藍藻毒素或過量養分在實踐中間環境突然釋放可能很重要。然而,本研究中用于改性天然土壤的殼聚糖可能對某些細菌(包括藍藻物種)具有抗菌活性。在實踐中應認真考慮絮凝劑的劑量。據報道,較高劑量的殼聚糖(例如>8 mg/L)可能導致銅綠微囊藻細胞裂解。本實驗中較低的殼聚糖劑量(3 mg/L)與Miranda等人(2017)報道的相似,他們發現對微囊藻沒有不利影響。此外,在我們之前的研究中,殼聚糖結合天然土壤可以進一步降低單獨使用殼聚糖對水生生物產生的毒性風險。
沉降后銅綠微囊藻的活力變化。
用土壤覆蓋可以使沉降的銅綠微囊藻細胞處于黑暗環境中,這是影響光合速率的關鍵因素。在本研究中,銅綠微囊藻細胞的光合作用和呼吸速率在絮凝和覆蓋處理后受到嚴重阻礙,這可能引發藻細胞分解。銅綠微囊藻細胞的光合作用和呼吸作用在F-no capping組中受到抑制,這反映在O2呼吸變化率顯著低于對照組。然而,在F-capping組中觀察到顯著較低的光合作用和呼吸速率,這表明通過F-capping處理產生了更高的光抑制效應。上述結果證實了絮凝-覆蓋處理可以加速藻華消亡的假設。應當指出,其他細菌(如異養細菌)對藻細胞呼吸的干擾應在進一步研究中加以探討。
在我們的實驗中,處理方式和溫度之間的交互作用可能顯著影響沉積的銅綠微囊藻細胞的光合作用和呼吸速率(P<0.05)。溫度是自然水體中藍藻生命活動的關鍵因素。在本研究中,設定了三個溫度(8°C、25°C和35°C)來模擬中國太湖春季、初夏和仲夏的實際溫度。優勢藍藻在春季快速生長,水華每年夏季在太湖發生。目前的結果表明,沉積的藻生物量在較低溫度下傾向于耐受弱光,這反映在對照組的正常形態、光合作用和呼吸作用上。類似地,Ma等人(2016)發現,大多數藍藻在低于12.5°C時沉入沉積層并保持休眠狀態(靜止細胞)。這些沉積的藻細胞在溫度升高時可能返回水柱,成為水華形成的潛在來源。較高的溫度刺激了銅綠微囊藻細胞的生長,這反映在35°C下對照組葉綠素a在15天前的增加速率比25°C下更快更高。F-capping系統中的O2消耗也隨著溫度升高而增加,并且在培養60天后,在25°C,特別是在35°C下,O2呼吸變為負值,表明較高的溫度加速了覆蓋層下掩埋藻華的呼吸速率。
水下植被對藻類氮的同化作用。
湖泊中營養循環主要發生在沉積物中,藻類沉降對生物地球化學過程具有顯著影響。藻華分解會直接釋放營養物質,可能導致沉積物和水體中營養成分循環的變化。39-41本研究發現,沉積的銅綠微囊藻會向沉積物釋放氮元素,隨后被脆皮藻吸收(圖4和圖5)。未被計算在內的15N可能源于底棲擾動和礦化作用。42含15N的有機氮可能通過硝化和反硝化反應轉化為無機態,最終以氣態形式凈流失。43未被計算的15N另一部分可能源自上覆水域釋放的營養物質(圖5)。此外,溫度升高會促進沉積物中微囊藻釋放更多氮元素,這與已有研究發現相吻合——營養循環速率會隨著藻華增加和溫度升高而加快。39大型水生植物對營養物質的吸收在緩解湖泊植被沉積物內部營養負荷方面起著關鍵作用。5本研究中檢測到脆皮藻生物量中存在過量15N,表明絮凝封蓋處理可將藻華釋放的營養物質轉移至沉水大型水生植物的生長環境中。因此,可以有效減少藻類釋放到水體中的過量氮(圖4-6)。這種方法在先前研究中已被廣泛采用28,44,用于恢復淺水區以沉水植物為主的健康生態系統。本研究發現,在8℃和25℃條件下,δ15N的快速吸收主要發生在實驗開始的前10天(圖4),這與普通蘆葦對標記銨和硝酸鹽的快速吸收現象一致45。此外,沉水植物對氮的同化作用可通過直接從水體中吸收氮來實現46,這可能是植被組中未檢測到的標記氮含量出現部分下降的原因(圖5)。較高溫度可能促進微囊藻來源的氮被脆皮藻同化(圖6)。這種現象的發生可能
部分原因在于沉積藻類生物質的分解和沉積物中有機氮的礦化過程,這些過程在較高溫度下會加速,從而為脆皮藻(P.crispus)提供更豐富的營養來源。此外,溫帶湖泊中的水生植物大多在春季至仲夏期間生長,這與我們的研究結果一致——脆皮藻在25°C時的生長速率是8°C時的兩倍(圖6)。生長速率顯著影響脆皮藻對δ15N的吸收,正如25°C條件下測得的δ15N‰值比8°C時高出五倍所反映的那樣。后續研究應重點關注脆皮藻生長后,與之相關的沉積藻華礦化速率。
湖泊修復的啟示。
通常情況下,在受人為富營養化影響的湖泊中,藻類向沉水植物的主導地位轉變,由于藻類生物量持續過度生長,自然條件下往往難以實現。即便在減少外部營養輸入后,要將這類湖泊從已形成的藻華狀態恢復到以沉水植物為主導的理想狀態,仍需大量人工干預。因此,許多湖內地球工程方法被廣泛采用,作為環保、高效且經濟的加速清除水體藻華的手段。19,20,28底水透明度和溶解氧濃度的改善得益于改良黏土/土壤技術的應用,19,28這種技術能促進(例如為植物發芽和生長創造特定周期)沉水植物的重建。本研究展示的絮凝封蓋法不僅能消除藻類生物量,還能加速其降解。隨后,腐爛藻類生物量釋放的營養物質可被沉水植被利用。高溫環境加速了藻類分解和融入植物生物量的過程,這表明在藻華期采用這種地球工程方法并結合播種大型水生植物,能夠通過藻類與沉水植被的生長季重疊(尤其在溫帶湖泊中),有效促進生態系統的這種轉變。不過,仍需通過試點田間實驗來驗證這種湖內地球工程方法的潛在效果——既能控制藻華現象,又能推動湖泊生態系統從藻類主導狀態向大型水生植物主導狀態的生態轉型。
