熱線:021-66110810,66110819
手機:13564362870
熱線:021-66110810,66110819
手機:13564362870
研究簡介:本文研究了毛竹(Phyllostachys edulis)枝條中縮短血紅蛋白(PeTrHb)在慢性缺氧適應中的調控作用。毛竹枝條的快速生長依賴于高效的能量代謝,但其密集的韌皮部結構和高耗氧量導致內部環境長期處于慢性缺氧狀態。本研究通過測量毛竹枝條不同生長階段的氧濃度、呼吸代謝,并結合RNA測序分析缺氧響應基因表達譜,揭示了PeTrHb在維持缺氧穩態中的潛在作用。研究發現,毛竹枝條內部氧水平(0.68%–3.60%[v/v])顯著低于土壤水中的氧水平(17.48%[v/v]),且基部低氧區域(富含分生組織的區域)與高PeTrHb1表達區域重疊。在空氣處理下,有氧呼吸僅占38%–47%,而在氮氣脅迫下,乙醇濃度并未顯著增加,表明毛竹枝條可能通過非乙醇發酵途徑維持能量穩態。PeTrHb1/2展現出極高的氧親和力(Ka=10^8–10^9 M^-1),其核質定位特征表明它們可能通過結合游離氧分子來限制線粒體的氧可用性。此外,通過在毛竹和瞬時轉化的煙草葉片中驗證,證實了PeTrHb1在維持缺氧下細胞活性中的關鍵作用。研究揭示了毛竹枝條在生長發育過程中存在的慢性缺氧現象,并提出了PeTrHb1可能通過調節氧分配來維持缺氧微環境的機制,這一機制可能避免了乙醇發酵的有毒積累,并為能量代謝提供了新的途徑。這些發現為未來分析毛竹快速生長與缺氧耐受之間的關系提供了參考,并為培育耐低氧脅迫的作物提供了潛在的靶點。
Unisense微電極系統的應用
Unisense微電極被用于測量毛竹枝條不同部位的溶解氧濃度。將針型氧微電極從頂端分生組織向下插入1cm處,測量溶解氧濃度。將微電極水平插入第五節間中心1cm處,進行溶解氧濃度的測量。在剝去鞘后,將傳感器水平穿透鞘基部區域,測量溶解氧濃度。通過精確測量毛竹枝條不同部位的氧濃度,為研究毛竹枝條內部的氧分布情況提供了直接數據,從而證實了毛竹枝條內部存在持續的低氧微環境,為后續研究毛竹枝條的慢性缺氧適應機制奠定了基礎。有助于了解毛竹枝條內部氧濃度與外部環境(如土壤水和枝條鞘)氧濃度的差異,以及不同生長階段氧濃度的變化情況,進而揭示了毛竹枝條內部氧濃度的分布特點和變化規律。
實驗結果
本研究揭示了毛竹枝條通過PeTrHb介導的潛在協調機制,該機制可能調和了快速生長期間的慢性缺氧與高能量需求之間的矛盾。盡管竹枝通過兩種途徑獲取氧氣,即竹鞘的氣孔氣體交換和根部介導的土壤溶解氧吸收,但由于細胞密度高、呼吸作用旺盛導致的氧梯度,分生組織卻發展出了持續的低氧狀態。高度表達的PeTrHb1可能會結合游離氧分子,降低細胞質中的氧濃度,從而限制線粒體的有氧呼吸。同時,細胞核中的PeTrHb1可能會參與調節代謝相關基因的表達,促使細胞依賴非乙醇發酵途徑來維持能量供應。這種“限氧-代謝適應”策略可能有助于支持竹枝分生組織中持續的細胞分裂和伸長。這些發現為快速生長植物在缺氧條件下PeTrHb介導的能量代謝調控提供了新的見解,同時也為培育抗低氧脅迫(如水淹)的作物提供了分子靶點。
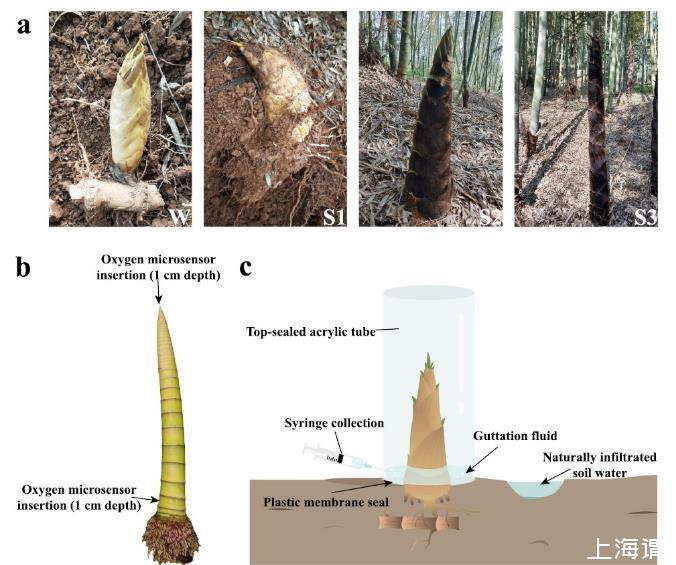
圖1、毛竹筍采樣和氧氣濃度測量示意圖.圖a顯示研究的四個生長階段:冬筍(W)、出土前春筍(S1)、50cm春筍(S2)和150cm高春筍(S3)。圖b展示氧氣測量方法:在頂端分生組織向下1cm處(頂端測量)和從基部向上第五節的中心水平插入1cm(基部測量)進行溶解氧濃度測定。圖c顯示吐水收集方法:用注射器收集吐水,并分析自然滲透土壤水的溶解氧濃度。
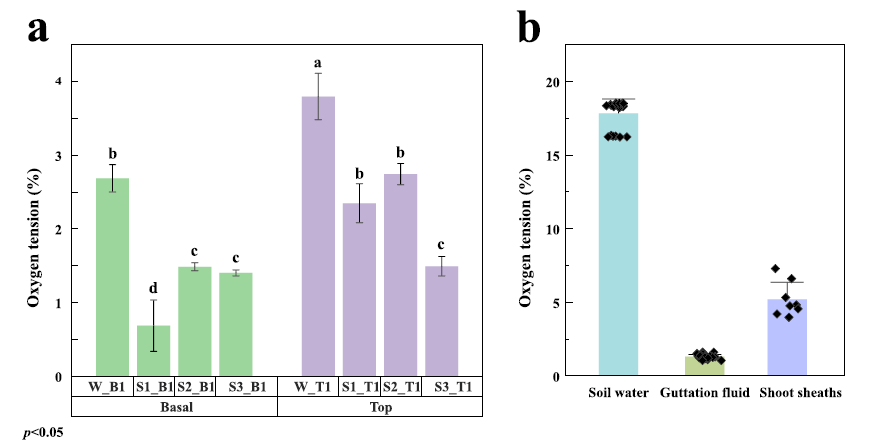
圖2、毛竹筍、土壤水、吐水和籜中的溶解氧濃度。圖a顯示不同生長階段竹筍基部和頂端區域的氧氣水平,字母不同表示Tukey檢驗顯著差異(p<0.05)。圖b比較了土壤水、吐水和籜中的氧氣濃度,顯示竹筍內部氧氣濃度(0.68-3.60%[v/v])顯著低于土壤水(17.48%[v/v])。
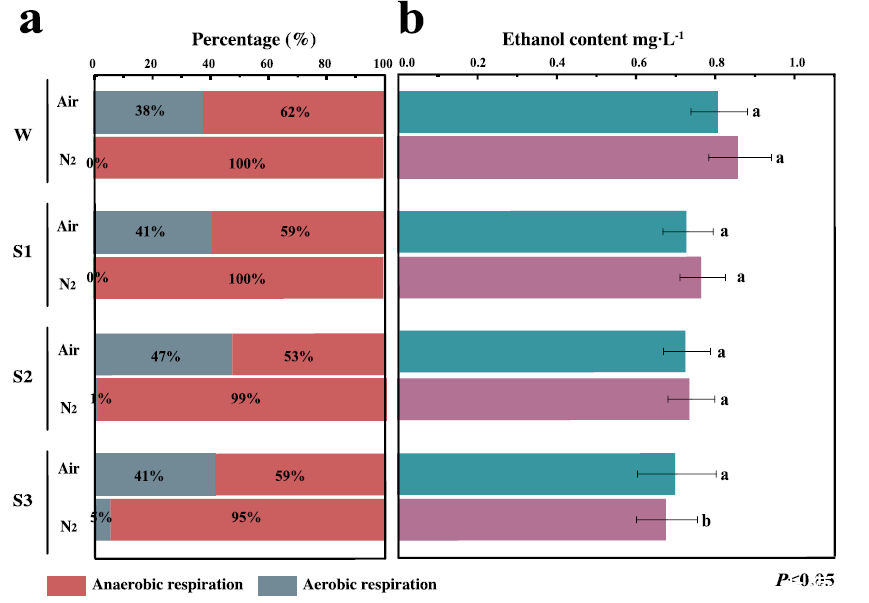
圖3、空氣和氮氣處理下毛竹筍(W-S3)有氧/無氧呼吸比例及乙醇濃度分析。圖中展示了呼吸代謝特征。(a)在空氣(Air)和氮氣(N2)處理條件下,莖中的有氧呼吸和無氧呼吸比例(W-S3)。(b)不同處理條件下莖組織中的乙醇濃度。
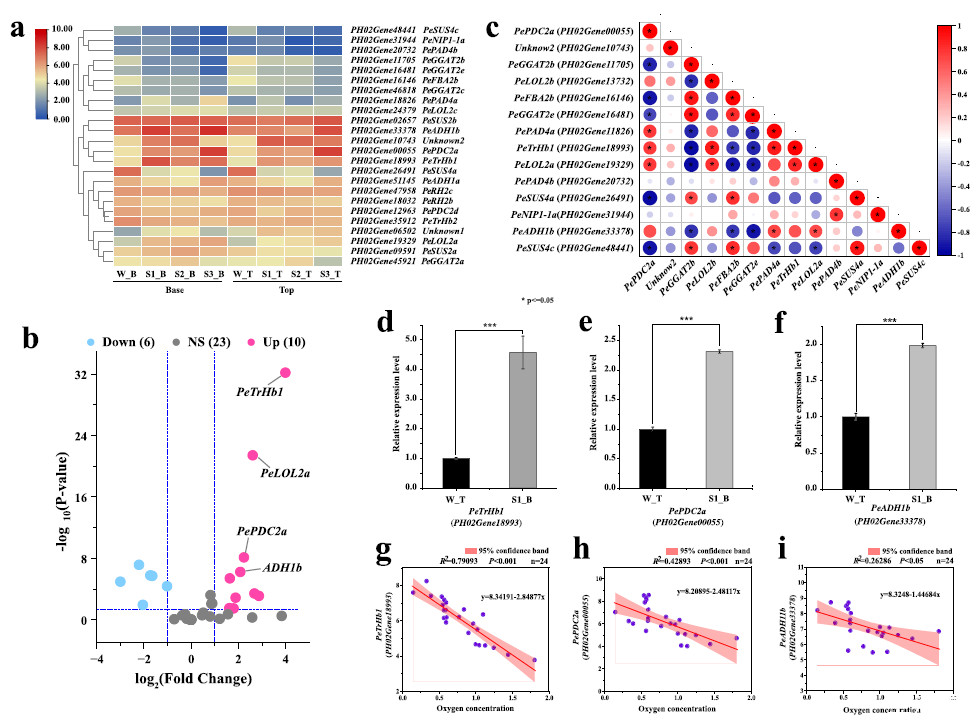
圖4、對毛竹嫩芽中缺氧響應基因的分析(a)GO:0001666 基因集表達的熱圖分析。樣本代碼:W_B(W 階段 - 基本型)、S1_B(S1 階段 - 基本型)、S2_B(S2 階段 - 基本型)、S3_B(S3 階段 - 基本型);W_T(W 階段 - 頂部)、S1_T(S1 階段 - 頂部)、S2_T(S2 階段 - 頂部)、S3_T(S3 階段 - 頂部)。(b)在 W-S3 階段的不同區域中 GO:0001666 基因集中差異表達基因的表達相關性分析。FPKM < 1 的基因被排除。(c)W_T(對照)與 S1_B 之間 GO:0001666 基因集中差異表達基因的火山圖。下:顯著下調的基因;NS:無顯著差異的基因;上:顯著上調的基因(p < 0.05),|倍數變化| > 2。(d)毛竹嫩芽中 PeTrHb1 的相對表達水平。(e)W_T 和 S1_B 中 PePDC2a(PH02Gene00055)的相對表達水平。(f)W_T 和 S1_B 中 PeADH1b(PH02Gene33378)的相對表達水平。(g)在毛竹莖中,氧濃度與 PeTrHb1(PH02Gene18993)表達量之間的線性回歸關系(n = 24,R2 = 0.79093,p < 0.001)。紅色陰影表示 95% 的置信區間。(h)在毛竹莖中,氧濃度與 PePDC2a 表達量之間的線性回歸關系(n = 24,R2 = 0.42893,p < 0.001)。(i)在毛竹莖中,氧濃度與 PeADH1b 表達量之間的線性回歸關系。
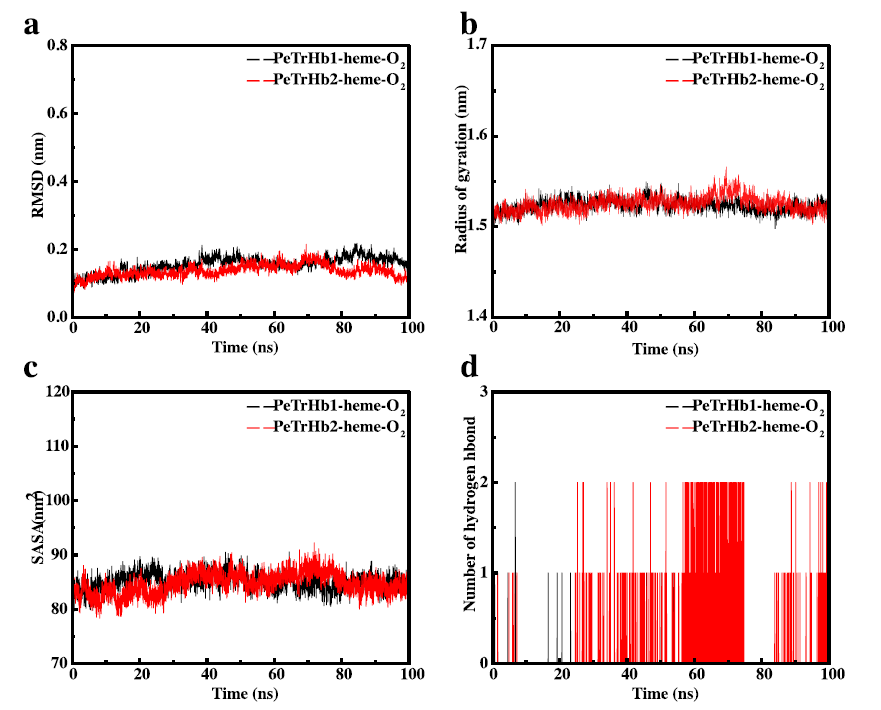
圖5、對PeTrHb、血紅蛋白和O2復合物的動力學模擬結果。(a)PeTrHb-血紅素-O2復合物系統的RMSD計算結果。(b)PeTrHb-血紅素-O2復合物系統的Rg計算結果。(c)PeTrHb-血紅素-O2復合物系統溶劑可及性SASA計算結果。(d)PeTrHb-血紅素-O2復合物系統中氧分子與氨基酸殘基之間氫鍵數量的計算結果。
結論與展望
毛竹筍的快速生長依賴于高效的能量代謝,但其致密的韌皮部結構和高耗氧量導致了慢性低氧環境。本研究測量了不同生長階段竹筍的氧氣濃度、呼吸代謝,并通過RNA-seq分析了低氧響應基因表達譜。結合分子動力學模擬、亞細胞定位和低氧脅迫實驗,我們鑒定了一個毛竹截短血紅蛋白(PeTrHb)并研究了其在維持低氧穩態中的潛在作用。結果顯示,竹筍內部氧濃度(0.68-3.60%[v/v])顯著低于土壤水(17.48%[v/v]),基礎低氧區(富含分生組織的區域)與PeTrHb1高表達區重疊。在空氣處理下,有氧呼吸比例僅為38-47%,但在氮氣脅迫下乙醇濃度未顯著增加,這些發現表明竹筍可能通過非乙醇發酵途徑維持能量穩態;PeTrHb1/2表現出超高的氧親和力(Ka=10?-10?M?1),其核質定位特征表明它們可能通過結合游離氧分子來限制線粒體氧可用性。最后,在毛竹和瞬時轉化的煙草葉片中驗證了PeTrHb1在維持低氧條件下細胞活力的關鍵作用。unisense微電極精確測量毛竹枝條不同部位的氧濃度,為研究毛竹枝條內部的氧分布情況提供了直接數據,從而證實了毛竹枝條內部存在持續的低氧微環境,為后續研究毛竹枝條的慢性缺氧適應機制奠定了基礎。本研究確定了毛竹筍生長發育過程中的慢性低氧現象,并提出PeTrHb1可能參與調節氧分配以維持低氧微環境,這種機制可能避免乙醇發酵的毒性積累,并為能量代謝提供新途徑。這些發現為未來分析毛竹快速生長與低氧耐受性之間的關系提供了參考。
 相關新聞
相關新聞