熱線:021-66110810,66110819
手機:13564362870
熱線:021-66110810,66110819
手機:13564362870
不同的CcoN亞基在菌落發育的早期或晚期對競爭適應性是必需的
為了進一步測試CcoN4對生物膜生長的貢獻,我們進行了競爭實驗,其中△N4和其他突變體與野生型一起作為混合菌株生物膜生長。在每個實驗中,一個菌株用組成型表達的YFP標記,以便在菌落形成單位(CFUs)計數期間區分菌株。實驗在每種菌株上標記進行,以確認YFP表達不影響適應性(圖3-圖補充1A,B)。當在菌落生長3天后評估競爭適應性時(圖3A),ΔN4細胞顯示出劣勢,野生型以兩倍的優勢勝過ΔN4。這與在AN1AN2突變體中觀察到的劣勢相似,進一步表明孤兒亞基CcoN4在生物膜代謝中起重要作用。
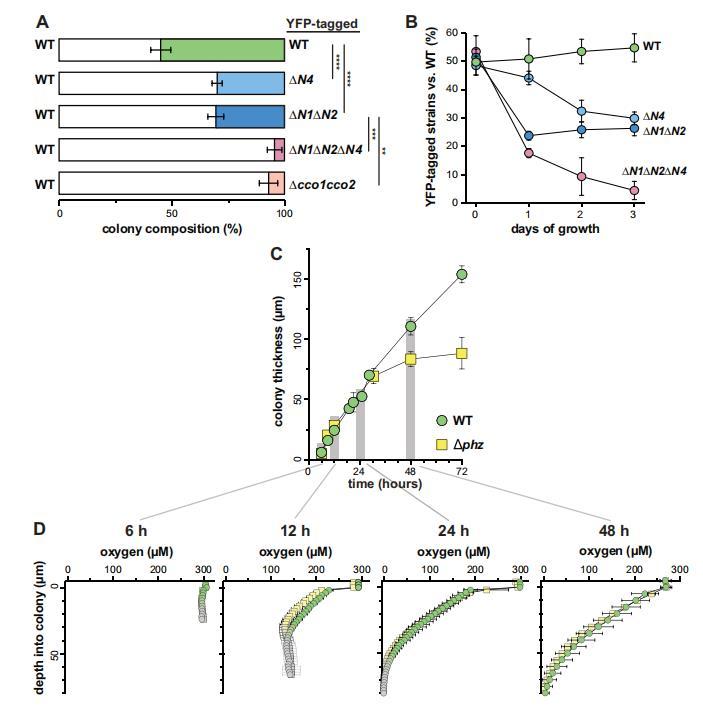
圖3 CcoN4在生物膜中具有競爭優勢,尤其是當O2成為限制條件時。(A)與WT在混合菌株生物膜中共同培養3天后,各種YFP標記的cco突變體的相對適應性。(B)顯示各種cco突變體與WT在混合菌株生物膜中共同培養3天的相對適應性的時間過程。圖中顯示的是WT與各種"標記"菌株(即設計為組成型表達YFP的菌株)共培養的實驗結果。(已標記的WT與未標記的突變體共培養的實驗結果見圖3-圖補充1)。誤差條代表生物三倍體的標準偏差。(C)通過薄片和DIC顯微鏡評估WT和Δphz的菌落生物膜在3天發育過程中的厚度變化。開始起皺后,測定基部(即皺紋之間的"谷")的厚度。(D)生物膜形成最初3天內選定時間點的菌落O2曲線。灰色點標記表示在菌落正下方的瓊脂中進行的測量。
DIC成像顯示野生型菌落薄切片在深度上存在形態變化,這可能是由于O2可用性降低所致(圖3-圖補充1C)。我們先前報道過,3天齡的PA14菌落生物膜在深處是缺氧的(Dietrich et al.,2013),并且在較薄的生物膜(如吩嗪缺失突變體Aphz形成的生物膜)中O2可用性通常更高。我們提出,在野生型生物膜中利用吩嗪作為電子受體使缺氧區的細胞生存成為可能,并促進菌落生長(Okegbe et al.,2014)。△N4突變體在競爭分析中相對較晚出現的表型(圖3B)使我們推測CcoN4可能在缺氧菌落亞區形成期間的生存中起作用,并且該區域可能在菌落生長1到2天之間出現。我們測量了野生型和Aphz生物膜在發育過程中特定時間點的O2濃度。
含CcoN4的異構體對PA14毒力具有獨特貢獻
我們之前已經證明,一個在生物膜特異性吩嗪生產中缺陷的突變體(其菌落形態也發生改變(Dietrich et al.,2008;2013))表現出毒力下降(Recinos et al.,2012)。我們和其他人提出,吩嗪可能通過作為電子受體在感染期間遇到的缺氧條件下平衡細胞內氧化還原狀態來促進毒力(Price-Whelan et al.,2006;Newman,2008;Dietrich et al.,2013)。由于CcoN4是野生型生物膜結構和呼吸作用所必需的(圖2A、C和圖5C),我們假設它也可能有助于毒力。為了驗證這一點,我們使用線蟲秀麗隱桿線蟲(Caenorhabditis elegans)作為宿主進行了毒力測定。研究表明,銅綠假單胞菌對秀麗隱桿線蟲具有致病性,并且緩慢致死模型模擬了細菌對秀麗隱桿線蟲的類似感染性殺死(Tan et al.,1999)。雖然△N1AN2以類似野生型的動力學殺死線蟲,但△N1ΔN2ΔN4和△cco1cco2相對于野生型PA14表現出同樣受損的殺死能力(圖6)。
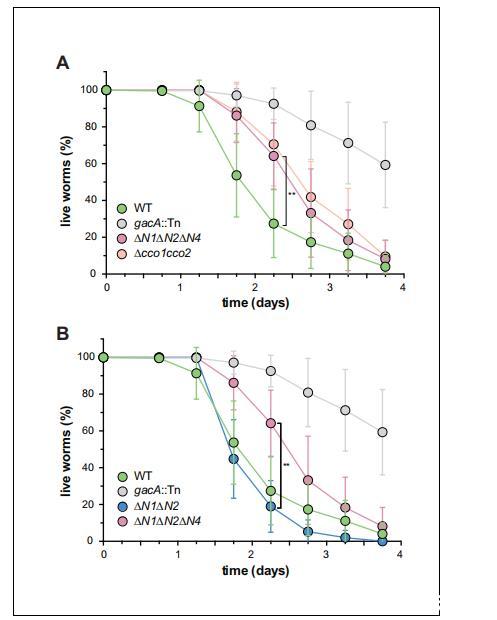
圖6含CcoN4的同工酶對PA14的毒力有獨特貢獻WT、gacA和各種cco突變株在線蟲秀麗隱桿線蟲中的緩慢殺滅動力學。暴露于WT PA14的秀麗隱桿線蟲群體在接觸該細菌4天后幾乎100%死亡,而缺乏GacA(一種控制銅綠假單胞菌毒力基因表達的調節因子)的突變體則顯示殺滅率下降,暴露后4天仍有約50%的蠕蟲存活。(A)與WT相比,ΔN1ΔN2ΔN4和Δcco1cco2的致病性明顯減弱。誤差條代表至少6個生物重復的標準偏差。暴露后2.25天,ΔN1ΔN2ΔN4殺死的elegans明顯少于WT(非配對雙尾t檢驗;p=0.0022)。(B)與WT相比,ΔN1ΔN2的致病性僅略有降低。暴露后2.25天,ΔN1ΔN2殺死的elegans明顯多于ΔN1ΔN2ΔN4(非配對雙尾t檢驗;p=0.003)。誤差條代表至少4個生物重復的標準偏差,每個重復的起始樣本量為30-35個蠕蟲。
討論
生物膜形成通常與宿主的定植和在宿主體內的持續存在相關,包括在囊性纖維化患者中觀察到的慢性肺部定植(Tolker-Nielsen,2014;Rybtke et al.,2015)。生物膜微環境內發現的條件與充分混合的液體培養物中的條件在電子供體和受體可用性方面是不同的。我們先前描述了由銅綠假單胞菌產生的電子穿梭抗生素吩嗪在生物膜特異性代謝中的作用。在本研究中,我們關注銅綠假單胞菌編碼cbb3型細胞色素氧化酶亞基的大量基因補體,并著手測試它們對生物膜中代謝電子流的貢獻。
銅綠假單胞菌基因組包含四個不同的ccoN同源物,編碼cbb3型氧化酶的催化亞基。其中只有兩個(ccoN1和ccoN2)與編碼活性cbb3型氧化酶另一關鍵成分的ccoO同源物共轉錄(圖1B)。然而,遺傳學研究已經證明,在銅綠假單胞菌PAO1中,所有四個版本的CcoN都可以在表達時與兩個CcoO同源物中的任何一個形成功能性復合物(Hirai et al.,2016)。在充分混合的液體培養物中,缺乏“孤兒”亞基的突變體沒有顯示出生長缺陷(圖2C)(Hirai et al.,2016)。因此,我們驚訝地發現,在菌落生物膜分析中,△N4突變體顯示出獨特的形態(圖2A,圖2-圖補充1A)。我們在研究細胞氧化還原平衡和感應的機制時廣泛使用了該分析,并注意到△N4的表型類似于電子穿梭和氧化還原信號缺陷突變體的表型(Dietrich et al.,2013;Okegbe et al.,2017)。
我們通過一系列分析表征了ΔN4突變對生物膜生理學的影響。在充分混合的液體培養物中,Dcco1cco2顯示出與DN1AN2相似的生長表型。雖然Hirai等人已證明在浮游生長條件下,野生型銅綠假單胞菌培養物確實形成含有CcoN4的Cco異質復合物,但我們的觀察表明,這些復合物在這些條件下對生長的貢獻不顯著。與此一致,在ΔN1ΔN2背景中刪除ccoN4對浮游生長沒有影響(圖2C)。然而,在基于生物膜的實驗中,我們發現單獨刪除N4就足以引起形態表型的改變(圖2A和圖2-圖補充1A),并且在DN1或AN1AN2背景中刪除N4深刻影響了生物膜生理學。這些實驗包括對菌落呼吸活性的量化,其中刪除CcoN4導致顯著降低(圖2B);生物膜共培養,其中CcoN4是競爭適應性所必需的(圖3A和B,圖3-圖補充1);氧化還原分析,顯示CcoN4可以促進吩嗪還原(圖5B,上圖);菌落厚度測量,顯示CcoN4是缺氧和缺氧區形成所必需的(圖5B,下圖);以及基質分析,顯示CcoN4有助于抑制Pel多糖的產生(圖5C)。在菌落薄切片中觀察到的cco1、cco2和ccoN4Q4在表達區域上的重疊(圖4)意味著CcoN4可以與Cco1和Cco2亞基形成跨越菌落深度的異質復合物,并以這些方式影響銅綠假單胞菌生物膜的生理學。

圖4 cco基因隨生物膜深度的不同而表達不同。左圖:從生長了3天的WT生物膜中制備的薄切片的代表性圖像。每個生物膜都在cco1、cco2或ccoN4Q4啟動子的控制下表達轉譯GFP報告基因。報告熒光顯示為綠色,并覆蓋在相應的DIC圖像上。右圖與左側圖像相對應的熒光值。各圖中均減去了含有gfp基因但無啟動子的菌株(空MCS對照)的熒光值。圖中還顯示了生長3天的WT生物膜的O2濃度隨深度變化的情況(開放圓圈)。
本研究報告的突變體表型和基因表達譜表明了CcoN4在生物膜特定環境下對O2和吩嗪還原的作用,并使我們能夠對其他CcoN亞基的作用得出結論。ccoN4Q4在整個生物膜深度上的表達表明含CcoN4的異構體可能參與有氧和缺氧區域的細胞色素c氧化(圖4)。這偏離了先前發表的觀察結果,即與充分通氣的液體培養物相比,這些基因在缺氧液體培養物中特異性誘導(Alvarez-Ortega and Harwood,2007)。因此,我們在菌落相對含氧的上部區域觀察到的ccoN4Q4表達可能是生物膜特有的。
AN4顯示出表明氧化還原應激的菌落形態,并且與野生型相比具有適應性劣勢(圖2A和圖3A,B,圖5B下圖,圖3-圖補充1)。然而,由于它在吩嗪還原方面沒有缺陷(圖5B,上圖),我們將其菌落形態和受損的適應性表型歸因于其在O2還原中提出的作用(Hirai et al.,2016)。類似地,ΔN1ΔN2顯示出相對于野生型的適應性降低(圖3A和B,圖3-圖補充1),同時顯示出與野生型相當的吩嗪還原(圖5B),這意味著其中一個或兩個亞基有助于生物膜中的O2還原。然而,當CcoN4與CcoN1和CcoN2一起被刪除時,所得菌株表現出嚴重的吩嗪還原缺陷,刪除兩個cco操縱子重現了這一表型(圖5B)。因此,我們的觀察表明cbb3型氧化酶在吩嗪還原中的作用,除了其已確立的O2還原作用之外,從而擴展了我們對它們在銅綠假單胞菌生理學和生存中整體貢獻的理解。
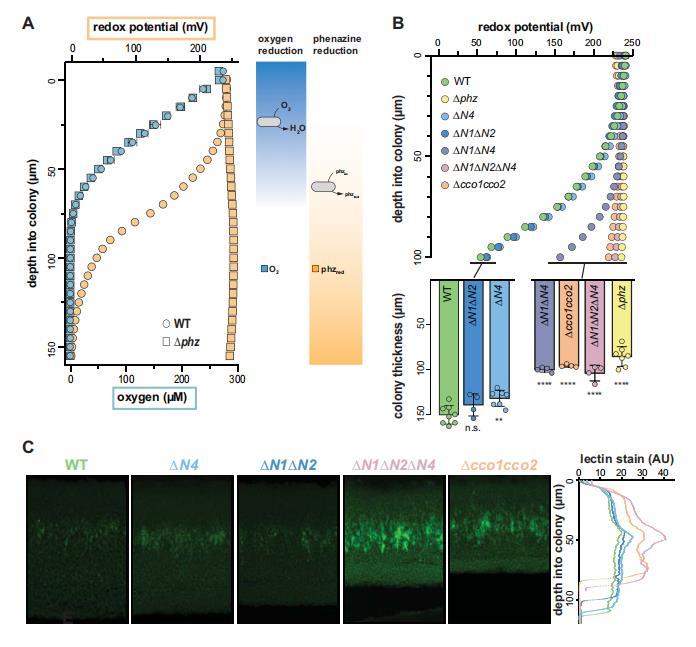
圖5 PA14 WT和突變體菌落生物膜中化學梯度和基質分布的特征。(A)左圖:生長兩天的WT和Δphz生物膜的氧氣濃度(藍色)和氧化還原電位(橙色)隨深度的變化。WT生物膜厚約150μm,而Δphz生物膜厚約80μm。對于O2曲線,誤差條代表生物三重樣的標準偏差。對于氧化還原曲線,數據代表至少5個生物重復。右圖:描述生物膜中O2和還原與氧化酚嗪類分布的模型。(B)頂部:生長2天的WT和各種突變生物膜的氧化還原電位隨深度的變化。至少5個生物重復的數據具有代表性。下圖所述菌株3天菌落生物膜的厚度。(C)左圖:WT和cco突變體生物膜的代表性薄切片,用凝集素染色并用熒光顯微鏡成像。生物膜在取樣前已生長2天。右圖凝集素染色信號強度的相對量化。左側面板中菌株名稱的顏色為繪制數據提供了一個關鍵,右側面板中的y軸為左側面板提供了一個刻度條。凝集素染色圖像和數值代表4個生物重復。
這里描述的結果可以啟發我們理解細胞如何在生物膜微環境內的不同條件下生存。先前的工作表明,丙酮酸發酵可以支持銅綠假單胞菌在缺氧條件下的生存(Eschbach et al.,2004),并且吩嗪促進這一過程(Glasser et al.,2014)。額外的研究表明,吩嗪還原是由銅綠假單胞菌黃素蛋白和脫氫酶偶然催化的(Glasser et al.,2017)。我們觀察到cbb3型細胞色素氧化酶,特別是含有CcoN1或CcoN4亞基的那些,是在缺氧生物膜亞區吩嗪還原所必需的(圖5B),這進一步將電子傳遞鏈與這些化合物的利用聯系起來。考慮到吩嗪在細胞色素bc1復合物和細胞色素氧化酶的生化研究歷史上作為介導劑的作用,這也很有趣(King,1963;Armstrong and Stewart-Tull,1971;Davidson et al.,1992)。基于這些早期工作,我們可以推測不同的CcoN亞基可能間接影響吩嗪還原,這可能發生在CcoO亞基的細胞色素c結合位點或電子傳遞鏈的其他地方,通過這些CcoN亞基對呼吸復合物的整體功能或穩定性產生影響。最終,在不同的生物膜深度或取決于電子供體可用性,吩嗪還原和吩嗪相關代謝的各種機制可能都是相關的。我們的結果表明,在菌落生物膜系統中,傳統上被認為是O2還原特異性的酶復合物可能有助于厭氧生存。
由于生物膜形成通常與宿主的定植和持續存在相關,我們測試了CcoN4是否有助于銅綠假單胞菌在秀麗隱桿線蟲中的致病性。類似于我們在生物膜分析中的觀察,我們發現Acco1cco2突變體比ΔN1ΔN2突變體表現出更嚴重的表型,表明孤兒亞基可以替代由cco1和cco2操縱子編碼的亞基。我們還發現在ΔN1ΔN2中刪除ccoN4導致了類似Dcco1cco2的表型,表明CcoN4是可以發揮這一作用的亞基(圖6)。在O2可用的宿主微環境中,含CcoN4的異構體可能有助于其還原。此外,在缺氧區域,含CcoN4的異構體可能促進吩嗪還原,從而實現細胞氧化還原平衡。這兩種功能都將有助于細菌在宿主體內的持續存在。cbb3型氧化酶對銅綠假單胞菌致病性的貢獻提出了干擾Cco酶功能的化合物可能成為這些感染有效療法的可能性。由于它們對細菌呼吸鏈的特異性,此類藥物將是極具吸引力的候選藥物,因此不會影響宿主的內源性呼吸酶。
我們發現一個孤兒cbb3型氧化酶亞基有助于生物膜生長,這進一步擴展了銅綠假單胞菌顯著呼吸靈活性的范圍。除了末端酶復合物水平的模塊性(例如利用aa3-型與cbb3-型氧化酶),銅綠假單胞菌呼吸鏈的活性還受到孤兒cbb3型催化亞基替代天然亞基的進一步影響。利用含CcoN4的異構體促進了吩嗪還原活性,并可能影響銅綠假單胞菌生物膜中的有氧呼吸。對于包含孤兒cbb3型催化亞基的特殊物種來說,這種精細的控制水平在覆蓋廣泛電子受體可用性范圍的環境(Cowley et al.,2015)中生長和生存可能特別有利。
 相關新聞
相關新聞