熱線:021-66110810,66110819
手機:13564362870
熱線:021-66110810,66110819
手機:13564362870
3.4吡啶脫氮過程中的硝化反硝化
將常用作硝化抑制劑的ATU加入到用PBR-1污泥接種的非原位間歇反應器中,比較其對吡啶和氮的去除性能。如圖3a-d所示,在最初的12小時內,吡啶可以在沒有ATU的情況下被完全去除,并且在該相中積累的NH4+占吡啶環中理論氮的10%-35%。吡啶完全去除后,NH4+-N濃度從7.4mg?L-1逐漸降低到0mg?L-1。但NO2--N和NO3--N濃度逐漸升高,分別達到1.1mg?L-1和6.3mg?L-1的峰值,然后下降到0mg?L-1。相應地,在36h時,TN液從最初的26.6mg?L-1明顯下降到0mg?L-1。在ATU存在下,12h對吡啶的去除率為81.5%,NH4+積累明顯,NH4+-N濃度峰值為11.5mg?L-1。隨后,在48h分批實驗結束時,NH4+-N濃度逐漸下降至1.1mg?L-1,但在整個過程中始終檢測不到亞硝酸鹽和硝酸鹽。相應地,TN液從最初的26.6mg·L-1下降到在36h時為2.3mg?L-1,始終高于無ATU時的水平。因此,可以推斷,因此,可以推斷氮的去除通過同化或硝化-反硝化與吡啶降解同時發生。無需外部碳源,通過反硝化作用可以去除NO2--N和NO3--N。根據李等人的研究,吡啶和外加硝酸鹽在COD/NO3-N為4和pH為7.5時可以有效去除。因此,可以推斷藻類分泌物和吡啶生物降解的中間體可以作為碳源支持脫氮。王等人指出,通過光合作用從CO2轉化的藻類分泌物可以作為細菌的碳源,而藻類可以通過有機物分解提供CO2和其他營養物質而從細菌中受益。在傳統的缺氧脫氮系統中,C/N比通常需要達到6或更高才能實現高脫氮效率(~90%)。然而,在ABA系統中,當C/N比約為4.3時,TN可以被完全去除。因此,在ABA系統中,PBR-1中的硝化-反硝化在吡啶和TN液去除中起著關鍵作用,與傳統的缺氧反硝化系統相比,其對外部碳源的要求較低。
電子受體,如硝酸鹽,可能通過反硝化活躍參與吡啶及其中間產物的生物降解。因此,向ABA系統中添加硝酸鹽以進一步確認反硝化參與吡啶去除(圖S6a-d)。在硝酸鹽添加的情況下,吡啶在9小時內可以完全去除,而在沒有硝酸鹽添加的情況下,吡啶去除效率僅為75.0%,表明硝酸鹽的添加顯著提高了吡啶的去除率。在24小時之前,NO2--N的逐漸積累和NO3--N的減少表明ABA系統中發生了可持續的硝化和反硝化。此外,吡啶生物降解產生的NH4+-N可以在40小時內完全去除,但硝酸鹽和亞硝酸鹽的去除速度較慢。反硝化的緩慢速度可能歸因于在吡啶完全去除后DO濃度急劇增加。因此,進一步驗證了反硝化參與了吡啶和氮去除的一部分。
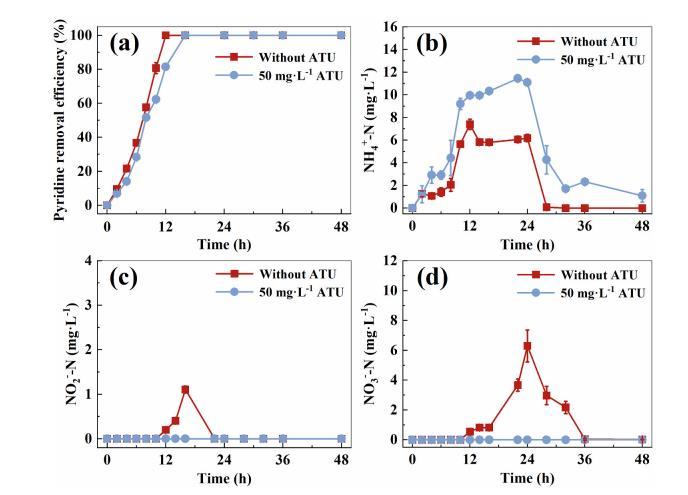
圖3吡啶去除效率(a)、NH4+-N濃度(b)、NO2--N濃度(c)和NO3--N濃度(d)隨硝化作用是否受到抑制(50 mg?L-1 ATU)的變化曲線。
不同的光照強度會影響光合活性和硝化作用。如圖4a-d所示,在270μmol·m-2·s-1的條件下,吡啶在8h內可被完全去除,而在180μmol·m-2·s-1和90μmol·m-2·s-1時吡啶去除效率分別為84.7%和57.7%,在90-270μmol·m-2·s-1范圍內,隨著光強的增加,可以加速對吡啶的去除。當光照強度為90μmol·m-2·s-1時NH4+-N積累峰值為7.4mg?L-1,在180μmol·m-2·s-1和270μmol·m-2·s-1時,NH4+-N積累峰值分別為3.5mg?L-1和3.7mg?L-1。加速吡啶的去除和減少NH4+-N的積累可能歸因于藻類通過光合作用增加了產氧量,然后氧增強了吡啶的降解和作為電子受體的硝化作用。當光照強度達到180μμmol·m-2·s-1及以上時,亞硝酸鹽和硝酸鹽沒有積累,這可能是由于硝化反硝化足夠快。因此,180μmol·m-2·s-1的光照強度足以實現吡啶和氮的有效去除。
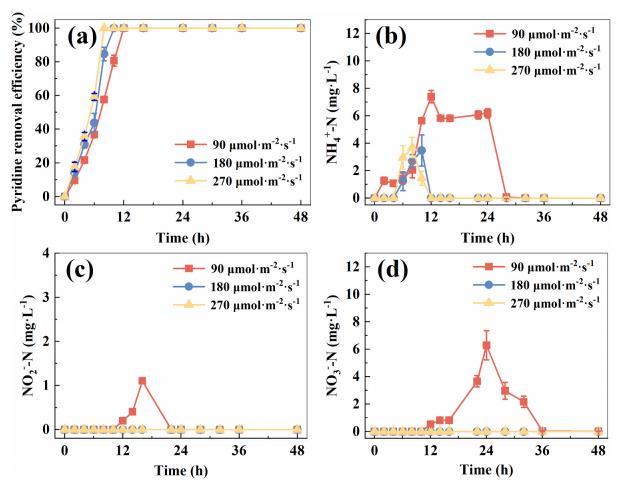
圖4不同光強下吡啶去除效率(a)、NH4+-N濃度(b)、NO2--N濃度(c)和NO3--N濃度(d)的圖譜。
為了研究額外碳源對ABA系統處理吡啶的影響,在分批實驗中,在不同時間加入乙酸鈉(NaAc)。如圖S7a-d所示,在0小時時,NaAc的加入降低了吡啶的降解速率。相應地,NH4+-N的積累在0h時隨著NaAc的加入而減少。這可能是由于NaAc與吡啶作為電子供體競爭氧氣,當NaAc和吡啶共存時,微生物更喜歡NaAc而不是吡啶。在之前的研究中也觀察到了類似的現象,在硝酸鹽存在的缺氧系統中,由于NaAc的添加,吡啶的生物降解受到了負面影響。此外,NH4+-N、NO2--N和NO3--N可以也可以在48小時內完全去除,這表明NaAc對ABA體系中亞硝酸鹽和硝酸鹽的去除沒有明顯的促進或抑制作用。因此,額外的碳源不能增強ABA體系中吡啶和氮的去除性能。
3.5同化和硝化反硝化對TN去除的貢獻
為了進一步研究脫氮途徑,根據方程(1)和方程(2)評估了同化和硝化反硝化對TN液去除的貢獻。如圖5a所示,在90μmol·m-2·s-1的光照強度下,硝化-反硝化和同化對TN液去除的貢獻率分別為40.9%和59.1%。然而,隨著光照強度增加到180μmol·m-2·s-1和270μmol·m-2·s-1,硝化反硝化對TN液去除的貢獻率分別提高到79.3%和75.5%,遠高于90μmol·m-2·s-1時的貢獻率。相應地,在180μmol·m-2·s-1和270μmol·m-2·s-1的光照強度下,同化對TN液去除的貢獻率分別明顯降低到20.7%和24.5%。當ATU抑制硝化作用時,TN液的去除率低至91.3%,明顯低于沒有ATU時的100%,證實了硝化反硝化在TN液去除中具有關鍵作用(圖5b)。如圖5c所示,考慮到在容易生物降解的有機物存在的情況下,自養微生物(如硝化物種)的生長將受到限制,硝化-反硝化作用對TN液去除的貢獻率隨著NaAc的加入而降低,這可能是由于硝化作用受到抑制。基于以上結果,可以推斷硝化反硝化作用在ABA系統中具有優先的脫氮作用。當氧氣產量充足時,硝化作用發生得很好,從而通過硝化反硝化作用。Vargas等人發現硝化作用和藻類吸收對氨的去除率分別為60%和40%,這與本研究一致。
對ABA系統的產氣過程進行了分析,進一步研究了附加碳源和硝酸鹽的影響,驗證了其脫氮過程。如圖S8所示,ABA系統產生的氣體由64.8%的O2和35.2%的N2組成,總氣體生成201μmol·L-1。添加NaAc(60.1%O2和39.9%N2)后,氣體生成量略微增加到212μmol·L-1(圖S7a-d)。然而,隨著NaNO3的加入,氣體生成顯著增加到660μmol·L-1(80.3%的O2和19.7%的N2)。反硝化作用參與吡啶及其中間體的生物降解可能是通過節省氧氣消耗來增加O2的原因。此外,氧氣的增加也可歸因于通過添加硝酸鹽作為氮源改善了藻類的生長和光合作用活性。本研究中產生的氣體僅為O2和N2,表明ABA系統中溫室氣體(如CO2和N2O)的排放量較低,這與其他ABS一致系統。
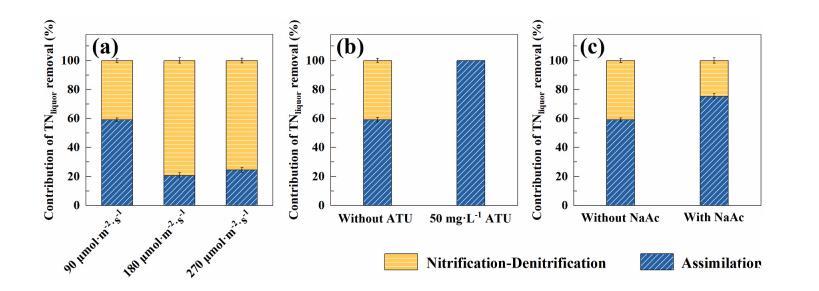
圖5對TN去除的貢獻不同光強度下去除(a)硝化抑制(50 mg?L-1 ATU)或無硝化抑制(b)和添加NaAc(c)
 相關新聞
相關新聞