熱線:021-66110810,66110819
手機:13564362870
熱線:021-66110810,66110819
手機:13564362870
3結果
3.1肉湯中氧飽和度對洛伐他汀生物合成的影響
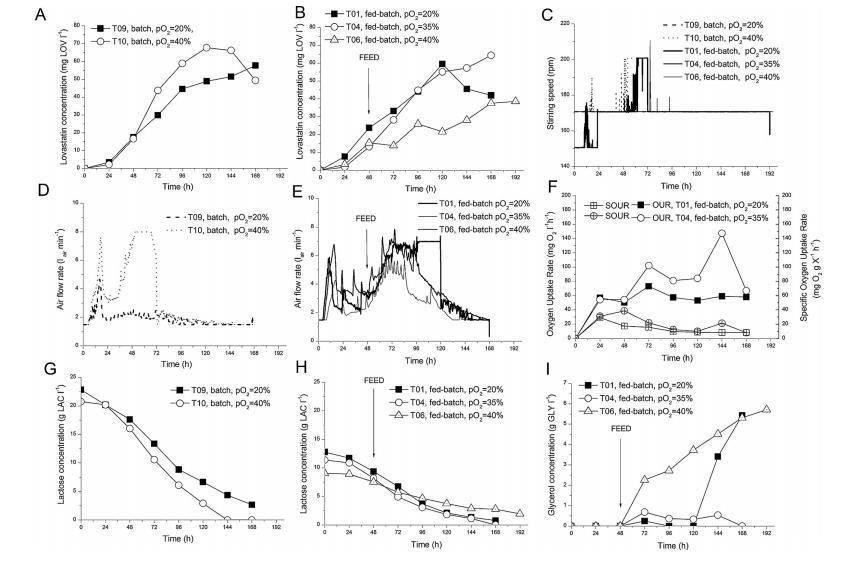
圖1、分批和連續分批補料實驗中培養基成分和工藝參數的時間變化,以顯示氧飽和度對洛伐他汀生物合成的影響:(A)分批運行中的洛伐他汀濃度,(B)連續分批補料運行中的洛伐他汀濃度,(C)攪拌速度,(D)間歇運行中的空氣流速,(E)連續分批補料運行中的空氣流速、(F)連續分批補料運行中的氧氣吸收率(OUR)和比氧氣吸收率(SOUR);(G)分批運行中的乳糖濃度;(H)連續分批補料運行中的乳糖濃度;(I)T01、T04、T06、T09和T10試驗中獲得的連續分批補料運行中的甘油濃度。
氧氣是影響土霉菌中洛伐他汀形成的最重要因素之一。它有幾個作用。首先,洛伐他汀的形成需要分解代謝途徑中碳源的高強度氧化,以形成丙二酰CoA,丙二酰CoA是洛伐他汀非酮類合成酶(LNKS)延伸聚酮鏈的單位。其次,洛伐他汀形成過程中所謂的后PKS階段需要氧氣,即在氧合酶催化下,由LNKS合成的中間體4a,5-二氫洛奈可林進行修飾。第三,真菌進行有氧生長,通過氧化途徑消耗碳底物以產生能量。
在第一組實驗中,研究了氧飽和度(pO2)對生物反應器中洛伐他汀生物合成的影響。圖1A和B顯示了在不同的pO2水平下,分批和連續分批補料生產時生產介質中洛伐他汀濃度的變化。土霉菌生產的洛伐他汀受pO2的影響,在分批補料工藝(運行T04)中,當pO2設為35%時,168小時的滴度為65mg LOV/l。在分批運行T10中,當氧飽和度為40%時,洛伐他汀的滴度在120小時時達到69mg LOV/l,但隨后由于乳糖的缺乏,形成的產物被重新利用(圖1F),在168小時時僅發現49毫克LOV/l。比較T09(pO2=20%)和T10(pO2=40%)兩個分批實驗,可以明顯看出,在72至120小時的運行期間,較高的氧飽和度有利于洛伐他汀的形成。另一方面,在pO2設為40%的甘油連續分批補料實驗(T06)中,洛伐他汀的滴度(約30毫克LOV/l)明顯低于pO2=40%的分批實驗,最終約6克GLY/l仍未在培養基中使用(圖1B和I)。
在這一組分批進行的實驗中,T04的結果最好(69mg LOV/l),其中pO2控制在35%,盡管肉湯中的洛伐他汀濃度直到96小時都低于T01(pO2=20%)。由于在所有實驗中肉湯中的氧飽和度都被控制在一個設定的水平,因此空氣流速(和攪拌速度,如果觀察到的話)的變化反映了土霉菌對氧氣的需求。圖1D和E顯示了選定運行的空氣流速。在分批補料運行(T01、T04和T06)中,補料開始時碳底物(甘油)供應的增加導致空氣流速增加。
值得注意的是,與運行T09相比,分批運行T10對氧氣的需求要高得多,在該運行中,洛伐他汀的生產率很高,氧飽和度被控制在40%的最高水平(圖1D)。在所有運行的早期滋養階段(12小時左右)以及分批補料運行T01和T06中(pO2控制在40%),氧氣吸收率非常高,以至于達到了空氣流速的上限,因此攪拌速度增加了(圖1C)。
在兩次分批補料實驗(T01和T04)中,氧氣需求量也用氧氣吸收率(OUR)和特定氧氣吸收率(SOUR)表示。這些速率隨時間的變化如圖1F所示。在設定了較高氧飽和度(35%)的實驗(T04)中,OUR和SOUR值均高于實驗T01。具體來說,晚期特異相中較高的OUR值與這一階段較高的洛伐他汀滴度相關(比較圖1B和F)。
在實驗T04、T06和T10中,為維持設定的pO2值而必須增加空氣流速和攪拌速度,這導致土霉菌從顆粒形態變為分散形態,這種變化影響了生物反應器中肉湯的流變特性。當出現分散形態時,生物反應器壁上的停滯區清晰可見,這表明在這些運行過程中肉湯具有剪切變稀的行為。這些停滯區在T04試驗中尤為明顯,在該試驗中,由于較高的剪切力(pO2設置為35%),在工藝開始時形成的蓬松(松散)顆粒后來轉變為分散形態。而在運行T01(pO2設為20%)時,流變學沒有發生這種變化:在工藝開始時形成的密度更大(更緊密)的顆粒在實驗后期沒有被破壞。在T01和T04試驗中,氧氣的對流傳質系數(kLa)值與土曲霉的形態(顆粒狀或分散狀)相吻合。在T01運行中,當顆粒占主導地位時,kLa在0.016到0.024s-1之間變化,而在T04運行中,當分散形態占主導地位時,kLa在72到120小時之間在0.0036到0.0085s-1之間變化。
最后,碳底物濃度隨時間的變化取決于pO2的控制水平(圖1G-I)。在氧氣較多(pO2=40%)的批次運行T10中,乳糖較早被完全耗盡,而在運行T09(pO2=20%)中,乳糖一直留在肉湯中,直到實驗結束(圖1G)。在分批補料實驗T01和T04中,無論設定的氧飽和度如何,乳糖的利用率都差不多,尤其是在開始補料之前(圖1H)。在分批進行的T06試驗(pO2=40%)中,乳糖的利用速度較慢,這可能是因為真菌形態發生了不利的變化,即由于機械應力導致顆粒和菌絲破壞。與此同時,甘油的利用速度也很慢,從開始補料起就一直在積累。這些因素綜合在一起,導致本實驗的洛伐他汀產量相對較低。而其他兩次分批進行的補料實驗則不同。在T04試驗中,所有添加的甘油幾乎全部被消耗,而在T01試驗中,甘油的濃度在試驗后期有所增加,達到約5.5g GLY/l。
總之,在分批生產過程中,較高的pO2值會加快碳底物(乳糖)的利用率,而眾所周知,這有利于洛伐他汀的形成。在分批式工藝中,40%的pO2水平是最佳的,但在補料式工藝中則不是。由于有更多的可用碳底物,在所有設定的pO2水平下,特別是在pO2=40%時(實驗T06),分批補料運行的洛伐他汀滴度應該會好得多。然而,這里的情況并非如此。在所有這些實驗中,除了真菌形態外,生物反應器中的所有工藝條件都是完全可控的。由于生物反應器中剪切應力的增加,真菌形態發生了不良變化,從最初的顆粒狀變成了分散的菌絲碎片。這些變化主要發生在pO2設置為35%或40%的運行過程中。從理論上講,在這些實驗中,菌絲體獲得更多的氧氣應該會提高洛伐他汀的滴度,但這并不像人們所期望的那樣明確。因此,我們進行了一項實驗,研究洛伐他汀滴度與顆粒內氧氣水平的關系。
3.2顆粒大小和結構對連續補料工藝中顆粒中氧氣分布的影響
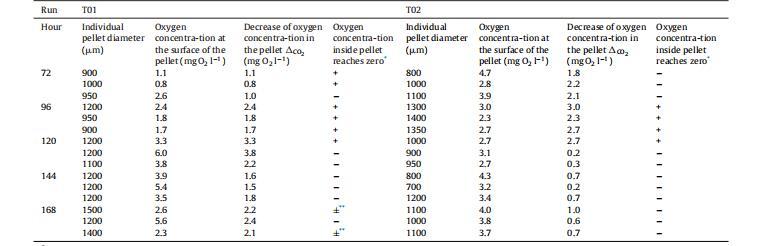
表1、實驗T01和T02(不同的培養前準備)的顆粒內氧氣濃度測量(每小時使用三個顆粒)。

圖2、生產培養基顆粒中的顆粒內氧濃度曲線:(A)T01運行72小時后;(B)T01運行144小時后;(C)T02運行72小時后和(D)T02運行144小時后;帶箭頭的垂直線代表顆粒表面;這樣,邊界層中的氧濃度梯度與顆粒內的梯度就區分開了。
在T01(見第3.1節)和T02兩項連續分批進行的實驗中,pO2控制在20%,以避免空氣流速和攪拌速度的大幅增加,從而將剪切應力降至最低。這樣可以在生物反應器中盡可能長時間地保持菌絲的顆粒結構,并測量顆粒內的氧氣濃度分布。此外,由于在預培養準備過程中采用了不同的方法從斜面上清洗孢子(見第2.1節),在這兩次實驗中獲得的顆粒大小略有不同,但結構卻大相徑庭。T02試驗中形成的顆粒稍小,結構松散、蓬松。還能觀察到一些分散的菌絲。而T01試驗形成的顆粒則更為緊湊。在這兩次實驗中,顆粒形態一直保持到168小時,而第3.1節中描述的其他實驗則沒有這種情況。因此,可以在靜態條件下測定顆粒內的氧氣濃度曲線(見第2.2節)。需要注意的是,這些測量過程中的傳質條件并不能準確反映生物反應器中的傳質條件。因此,根據顆粒表面的氧氣濃度和顆粒內部(即表面和中心之間)氧氣濃度的下降情況,對從T01和T02實驗中取出的顆粒進行了簡化比較。表1顯示了所有這些結果。
對于在T02試驗中獲得的結構松散的稍小顆粒,只有少數顆粒內部的氧濃度降為零。而在T01試驗中獲得的結構更緊湊的較大顆粒中,在不同培養時間測試的15個顆粒中有8個的氧氣濃度為零。此外,在T01試驗的顆粒中,從顆粒表面到中心的氧氣濃度(CO2)下降幅度(平均為1.9毫克O2/l)高于T02試驗的顆粒(平均為1.3毫克O2/l)。
圖2顯示了顆粒內氧氣濃度分布的示例。在運行T02的結構密度較低的顆粒中,氧氣深入顆粒內部。這種情況在72小時內尤為明顯,這對高效生產洛伐他汀至關重要。72小時時,通常是洛伐他汀生產率和乳糖利用率最高的時候,只有在T01運行的顆粒中,也就是在形態致密的顆粒中,氧氣濃度才降到零(圖2A和C)。當乳糖成為限制條件時,兩次運行的顆粒中的氧濃度再次上升(圖2B、D和圖3B),表明氧消耗速度減慢。在T02試驗中,氧濃度的增加高于T01試驗,這與肉湯中檢測到的乳糖含量較低有關(圖3B)。
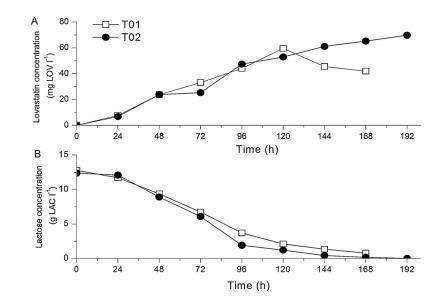
圖3、在不同的培養前準備和接種(T01和T02)實驗中,(A)洛伐他汀和(B)乳糖濃度的時間變化。
3.1顯示,通過提高肉湯中氧飽和度的設定水平,提高了洛伐他汀的生物合成。本節的結果表明,確保顆粒內的高氧濃度也有利于提高洛伐他汀的滴度(圖3A):168小時后,氧飽和度較高、顆粒較松散的T02試驗的洛伐他汀滴度比顆粒較緊湊的T01試驗高出25毫克LOV/l。
盡管稍小和較松散的顆粒對洛伐他汀滴度的增加作用不大,但我們的研究結果表明,控制真菌形態有利于土霉菌生產洛伐他汀。因此,我們采用了微顆粒強化培養技術(MPEC)。
