熱線:021-66110810,66110819
手機:13564362870
熱線:021-66110810,66110819
手機:13564362870
In-Situ Metatranscriptomic Analyses Reveal the Metabolic Flexibility of the Thermophilic Anoxygenic Photosynthetic Bacterium Chloroflexus aggregans in a Hot Spring Cyanobacteria-Dominated Microbial Mat
原位元轉錄組學分析揭示了嗜熱無氧光合細菌 Chloroflexus 聚集體在高溫 環境中的代謝靈活性
來源:Microorganisms 2021, 9, 652.
一、摘要概述
本研究通過原位宏轉錄組分析結合微尺度環境監測,揭示了嗜熱無氧光合細菌Chloroflexus aggregans在日本中房溫泉藍藻主導微生物墊中的代謝策略。核心發現包括:
晝夜代謝轉換:
日間高光高氧期(11:00):以光異養代謝為主(利用有機碳源),伴隨固碳基因低表達。
晨間微氧期(05:00):以化學混合營養代謝為主(同時利用H?、CO、有機碳),呼吸鏈基因(如氫酶、細胞色素氧化酶)高表達。
環境驅動機制:光照強度與O?濃度動態變化(通過Unisense微電極監測)直接調控基因轉錄模式。
生態意義:首次證實C. aggregans在自然環境中通過代謝靈活性適應晝夜環境波動,挑戰了傳統認為綠彎菌門為嚴格厭氧光合菌的認知。
二、研究目的
解析代謝策略:闡明C. aggregans在動態溫泉環境中的能量獲取與碳固定途徑。
驗證代謝靈活性假說:探究其如何協調光養、化養、自養與異養代謝以適應晝夜變化。
揭示生態角色:明確其在藍藻墊中的初級生產者功能及與共生微生物的互作關系。
三、研究思路
采用 "原位監測→轉錄組分析→代謝通路整合" 策略:
環境參數動態監測:
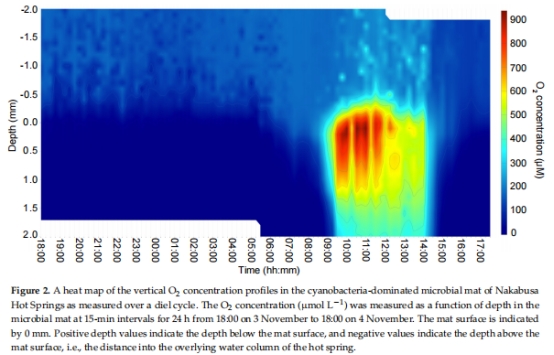
使用Unisense O?微電極(Clark型)每15分鐘監測微生物墊0–2 mm深度O?濃度剖面(圖2)。
同步記錄光照強度(PAR, 400–700 nm)。
時間序列采樣:
在24小時內(11個時間點)采集微生物墊樣品(圖1),提取總RNA。

宏轉錄組分析:
比對至C. aggregans參考基因組(NC_011831.1),分析能量代謝(光合、呼吸)、碳固定(3-羥基丙酸雙循環)、電子供體利用(H?、CO、硫化物)相關基因表達。
數據關聯:
將基因表達譜與O?/光照動態關聯,推斷主導代謝模式。
四、關鍵數據及研究意義
1. O?與光照動態(圖2)
數據來源:Unisense微電極連續24小時監測(圖2熱圖)。
結果:
正午(11:00):O?濃度 >900 μmol·L?1(超氧狀態),滲透深度 >2 mm。
清晨(05:00):O?濃度 <100 μmol·L?1(微氧狀態),僅存于表層。
意義:量化了微生物墊的氧化梯度,為解釋代謝轉換提供環境背景。
2. 光合相關基因表達(圖3)
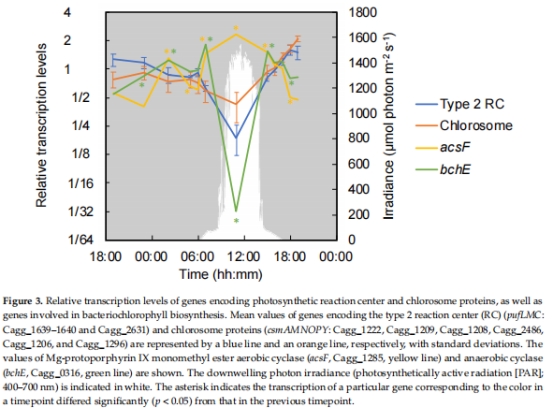
數據來源:光合反應中心(pufLMC)、綠小體蛋白(csmAMNOPY)基因。
結果:夜間高表達(厭氧期合成色素),日間受O?抑制。
意義:證實光合裝置合成受O?負調控,適應晝夜缺氧/富氧交替。
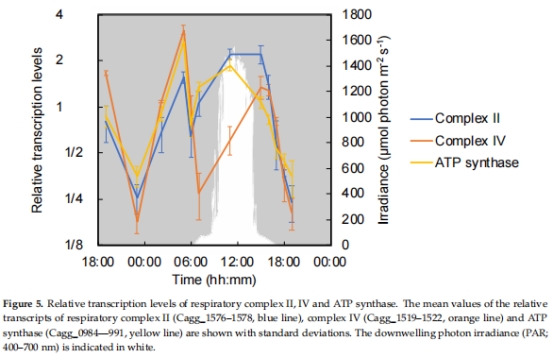
3. 呼吸與碳代謝基因表達(圖4–8)
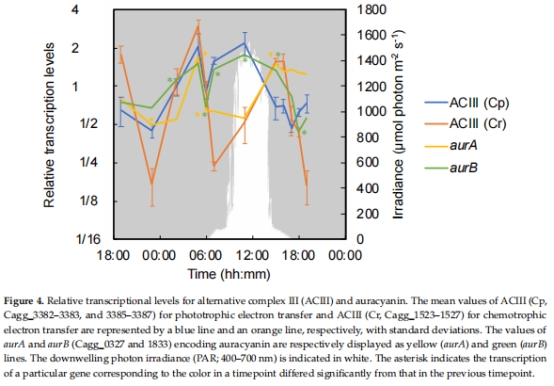
數據來源:
呼吸鏈:氫酶(hyd)、細胞色素氧化酶(cox)、ATP合酶(圖5)。
碳固定:3-羥基丙酸雙循環關鍵酶(malonyl-CoA reductase)(圖6)。
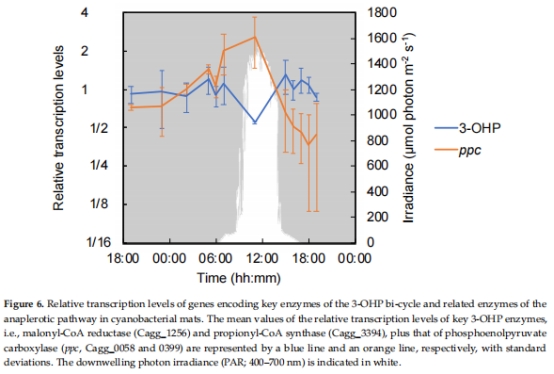
電子供體利用:H?酶(hyd)、CO脫氫酶(coxGSML)、硫化物:醌氧化還原酶(sqr)(圖7)。
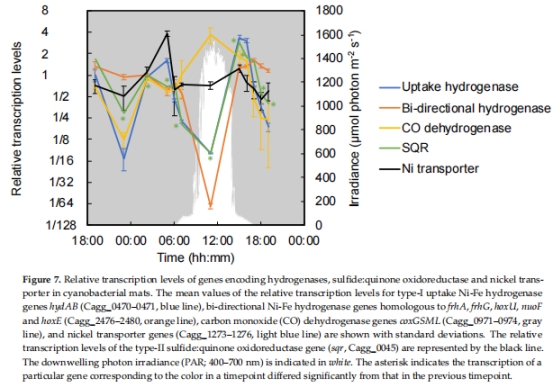
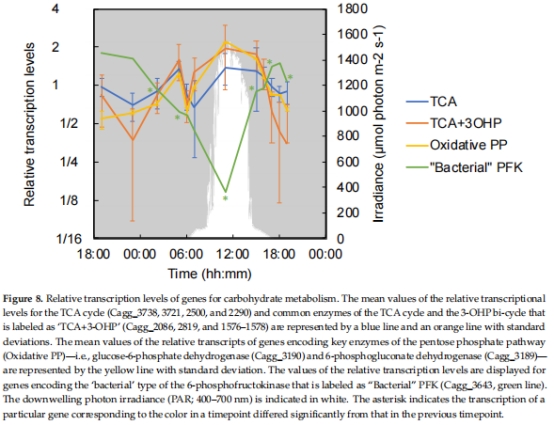
結果:
清晨(05:00):hyd、cox、TCA循環基因高表達,提示化學混合營養。
正午(11:00):糖轉運蛋白基因高表達,提示光異養主導。
意義:揭示代謝靈活性分子基礎,闡明C. aggregans如何利用不同電子供體(H?、CO、有機物)。
4. 晝夜代謝模式整合(圖9)
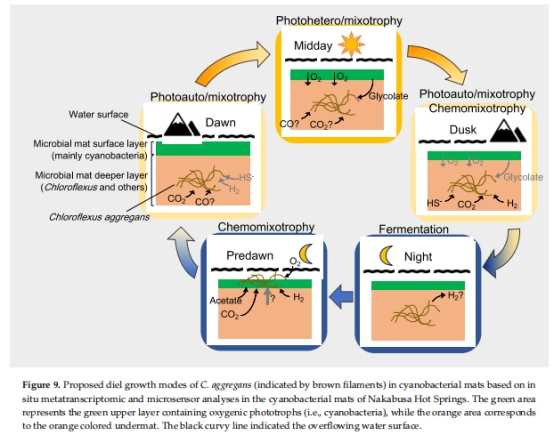
數據來源:綜合上述基因表達與環境參數。
結論模型:
日間:光異養(高光高氧)→ 光自養(低光微氧)。
夜間:發酵代謝 → 清晨:化學混合營養(微氧下利用H?/CO自養+有機物異養)。
五、丹麥Unisense電極數據的詳細解讀
1. 技術原理與實驗設計
原理:Clark電極通過O?在鉑陰極還原產生電流,靈敏度達μmol·L?1級(響應時間<0.5 s)。
設計:
垂直剖面監測(0–2 mm深度,100 μm分辨率)。
每15分鐘自動記錄剖面,持續24小時(圖2)。
2. 關鍵結果與生態意義
O?動態的代謝驅動:
正午超氧(>900 μmol·L?1):藍藻光合作用導致,抑制C. aggregans厭氧代謝基因(如bchE)。
清晨微氧(<100 μmol·L?1):支持C. aggregans呼吸代謝(hyd、cox高表達)。
空間異質性:
表層(0–0.5 mm):晝夜O?波動劇烈,驅動代謝快速轉換。
深層(>1 mm):持續缺氧,支持厭氧光合/發酵。
技術優勢:
高時空分辨率:捕捉O?梯度瞬態變化(如午后O?速降),關聯基因表達時移。
原位真實性:避免采樣擾動,真實反映微生物生存微環境。
3. 對代謝模型的驗證作用
證偽嚴格厭氧假說:檢測到晨間微量O?(<100 μmol·L?1),支持化學混合營養模式。
量化代謝窗口期:確定05:00和15:00為代謝轉換關鍵節點,指導轉錄組采樣設計。
揭示遷移行為:O?剖面暗示C. aggregans晨間向微氧表層遷移,趨氧性可能驅動代謝轉換。
六、結論
代謝靈活性:C. aggregans通過晝夜代謝轉換(光養→化養、自養→異養)適應動態環境。
生態功能:
日間:利用藍藻分泌物(如乙醛酸)光異養生長。
清晨:利用H?/CO化學自養,成為初級生產者(圖9)。
技術啟示:原位宏轉錄組+微傳感器聯用是解析復雜微生物群落功能的有效范式。
圖表示例說明
圖1:溫泉采樣點及微生物墊結構(藍藻層+綠彎菌層)。
圖2:O?濃度垂直剖面熱圖(Unisense數據)。
圖3:光合基因晝夜表達模式。
圖5:呼吸鏈基因表達與光照/O?關聯。
圖7:電子供體利用基因表達動態。
圖9:整合代謝模型(光養/化養轉換示意圖)。