熱線:021-66110810,66110819
手機(jī):13564362870
熱線:021-66110810,66110819
手機(jī):13564362870
Dissimilatory nitrate reduction by a freshwater cable bacterium
淡水電纜細(xì)菌的異化硝酸鹽還原
來源:The ISME Journal (2022) 16:50–57;
摘要核心內(nèi)容
本文首次證實(shí)淡水電纜細(xì)菌(Candidatus Electronema sp. GS)通過 異化硝酸鹽還原為銨(DNRA) 途徑還原硝酸鹽(圖3)。研究結(jié)合電化學(xué)電位測量(Unisense電極)、1?N同位素示蹤和轉(zhuǎn)錄組分析,發(fā)現(xiàn):
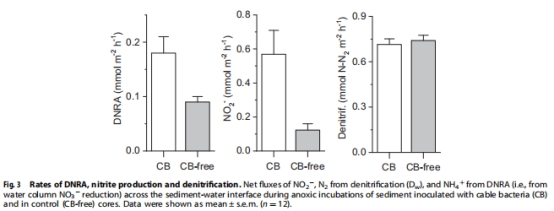
DNRA速率(89.7 ± 16 μmol NH?? m?2 h?1)和亞硝酸鹽產(chǎn)量(446 ± 73 μmol NO?? m?2 h?1)在電纜細(xì)菌富集沉積物中顯著高于對照組(圖3)。
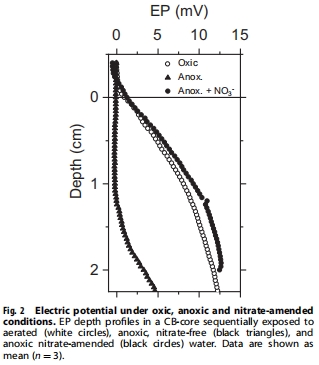
陰極硝酸鹽還原的電子通量(1.27 ± 0.06 mmol e? m?2 h?1)與DNRA代謝需求(1.61 ± 0.1 mmol e? m?2 h?1)匹配(圖2),證實(shí)DNRA由跨膜電子傳遞驅(qū)動。
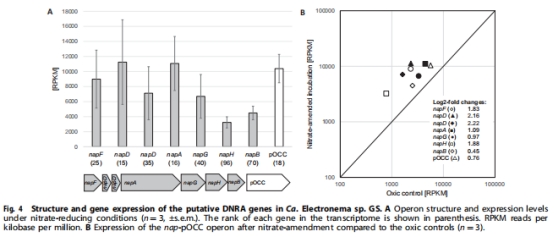
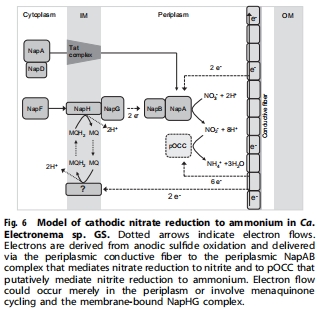
nap操縱子和假想八血紅素細(xì)胞色素c(pOCC)基因高表達(dá)(圖4),暗示其參與硝酸鹽→亞硝酸鹽→銨的轉(zhuǎn)化路徑(圖6)。
研究目的
驗(yàn)證假設(shè):探究電纜細(xì)菌是否通過DNRA途徑還原硝酸鹽(而非反硝化)。
解析機(jī)制:闡明DNRA的電子傳遞路徑與關(guān)鍵酶功能(如pOCC)。
進(jìn)化溯源:追溯硝酸鹽還原基因的水平轉(zhuǎn)移事件。
研究思路
采用 多尺度實(shí)驗(yàn)驗(yàn)證框架:
沉積物培養(yǎng):
熱滅活沉積物接種電纜細(xì)菌(CB組) vs. 未接種對照組(CB-free組)。
好氧預(yù)培養(yǎng)后切換至缺氧硝酸鹽環(huán)境(200 μM 1?NO??)。
電化學(xué)監(jiān)測:
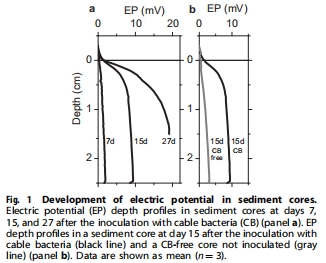
Unisense電位電極測量沉積物 電位梯度(EP)(圖1-2),計(jì)算電子通量。
1?N示蹤:
量化DNRA、亞硝酸鹽生成及反硝化速率(圖3)。
分子機(jī)制:
轉(zhuǎn)錄組分析 nap和 pOCC基因表達(dá)(圖4)。
系統(tǒng)發(fā)育樹重建基因進(jìn)化史(圖5)。
測量數(shù)據(jù)及其研究意義
1. 電位梯度(EP)與電子傳遞(機(jī)制尺度)
數(shù)據(jù)來源:
EP剖面:Unisense電極捕捉陰極區(qū)0.95 V m?1電位差(圖2),證實(shí)硝酸鹽驅(qū)動電子傳遞。
電子通量:計(jì)算陰極硝酸鹽還原電子流為1.27 ± 0.06 mmol e? m?2 h?1(圖2)。
研究意義:
首次量化DNRA的 跨膜電子需求,建立電子通量與DNRA速率的能量平衡(1.27 vs. 1.61 mmol e? m?2 h?1)。
排除反硝化途徑:反硝化速率在CB與對照組無差異(圖3),且N?O未激發(fā)電位響應(yīng)。
2. 氮轉(zhuǎn)化速率(過程尺度)
數(shù)據(jù)來源:
DNRA速率:CB組顯著高于對照組(↑100%,圖3)。
亞硝酸鹽生成:CB組達(dá)446 ± 73 μmol m?2 h?1(圖3)。
研究意義:
證實(shí)電纜細(xì)菌主導(dǎo) 硝酸鹽→亞硝酸鹽→銨 的還原路徑,增加沉積物氮滯留(非脫氮)。
3. 基因表達(dá)與進(jìn)化(分子尺度)
數(shù)據(jù)來源:
轉(zhuǎn)錄組:napFDDAGHB-pOCC基因簇高表達(dá)(RPKM排名前100,圖4A)。
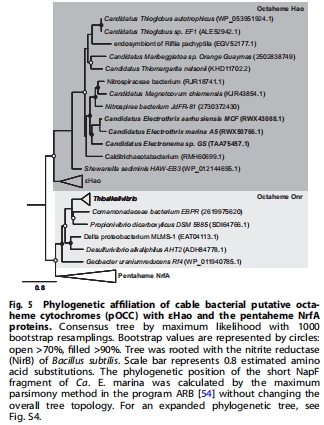
系統(tǒng)發(fā)育:nap和 pOCC基因水平轉(zhuǎn)移自α/β/γ-變形菌(圖5)。
研究意義:
pOCC基因定位DNRA途徑關(guān)鍵酶(圖6),但功能需純化驗(yàn)證。
進(jìn)化分析表明DNRA能力在淡水/海洋電纜細(xì)菌分化前獲得。
結(jié)論
生理機(jī)制:
電纜細(xì)菌通過 DNRA途徑 還原硝酸鹽至銨,電子由陽極硫化氫氧化經(jīng)導(dǎo)電纖維傳遞至陰極(圖6)。
陰極細(xì)胞 不通過氧化磷酸化產(chǎn)能,僅作電子"泄放口"。
生態(tài)意義:
DNRA增加生態(tài)系統(tǒng) 氮滯留(vs. 反硝化脫氮),可能影響河口氮循環(huán)平衡。
進(jìn)化啟示:
nap和 pOCC基因經(jīng)多次水平轉(zhuǎn)移獲得,淡水/海洋菌株共享祖先DNRA能力。
Unisense電極數(shù)據(jù)的專項(xiàng)解讀
技術(shù)優(yōu)勢
原位高分辨:
毫米級電位剖面(圖1-2)實(shí)時(shí)捕捉 生物電化學(xué)活動,避免沉積物擾動。
電子通量量化:
基于EP梯度計(jì)算陰極電子流(公式1),直接關(guān)聯(lián)DNRA代謝需求。
科學(xué)價(jià)值
機(jī)制驗(yàn)證:
電位響應(yīng)硝酸鹽添加(圖2)排除反硝化途徑,鎖定DNRA為唯一硝酸鹽還原路徑。
電子供需平衡(1.27 vs. 1.61 mmol e? m?2 h?1)為DNRA提供 直接能量證據(jù)。
模型構(gòu)建:
電極數(shù)據(jù)支撐 跨細(xì)胞電子傳遞模型(圖6),闡明電纜細(xì)菌"生物導(dǎo)線"功能。
研究意義
修正氮循環(huán)認(rèn)知:電纜細(xì)菌作為新型DNRA驅(qū)動者,增加沉積物氮滯留潛力。
技術(shù)示范:Unisense電極為沉積物生物電化學(xué)研究提供范式。
總結(jié):本研究通過Unisense電極的高精度電位監(jiān)測,首次揭示淡水電纜細(xì)菌以DNRA途徑耦合硫氧化與硝酸鹽還原,其陰極電子傳遞驅(qū)動的獨(dú)特代謝機(jī)制為環(huán)境生物電化學(xué)研究提供新視角。電極數(shù)據(jù)定量關(guān)聯(lián)電子通量與氮轉(zhuǎn)化,為復(fù)雜沉積物系統(tǒng)提供不可替代的解析能力。