熱線:021-66110810,66110819
手機:13564362870
熱線:021-66110810,66110819
手機:13564362870
Mitochondrial rewiring drives metabolic adaptation to NAD(H) shortage in triple negative breast cancer cells
線粒體重新布線驅動三陰性乳腺癌細胞對 NADH短缺的代謝適應
期刊:Neoplasia
來源:ScienceDirect (DOI: https://doi.org/10.1016/j.neo.2023.100903)
摘要核心內容
本研究探討三陰性乳腺癌(TNBC)細胞對NAMPT抑制劑FK866耐藥的代謝適應機制。通過建立FK866耐藥細胞模型(MDA-MB-231 RES),研究發現:
耐藥機制獨立于經典藥泵外排或補償性NAD?合成通路(如NMRK1),而依賴線粒體代謝重組。
線粒體功能增強:耐藥細胞線粒體備用呼吸能力(SRC)、線粒體質量(mtDNA/nDNA比例增加)及生物合成(PGC-1α/TFAM上調)顯著提升。
代謝底物依賴性改變:耐藥細胞更依賴丙酮酸和琥珀酸氧化供能,且線粒體丙酮酸載體MPC2的抑制可誘導敏感細胞產生類耐藥表型。
關鍵結論:持續NAD(H)短缺驅動線粒體重編程,通過增強氧化磷酸化(OXPHOS)和底物靈活性(丙酮酸→琥珀酸轉換)維持能量穩態,導致FK866耐藥。
研究目的
探究TNBC細胞對NAMPT抑制劑FK866耐藥的分子機制,重點關注線粒體代謝適應在抵抗NAD(H)短缺中的作用,為克服耐藥提供新靶點。
研究思路
模型建立:
通過長期遞增FK866暴露,構建MDA-MB-231耐藥細胞系(RES)。
耐藥機制初篩:
排除NAMPT突變、藥泵外排(ABCB1/ABCC1)及補償性NAD?合成通路(NMRK1沉默驗證)。
線粒體功能聚焦:
Seahorse分析糖酵解率(GlycoPER)和線粒體呼吸功能(OCR)。
評估線粒體質量(mtDNA/nDNA比例、MitoTracker染色)、生物合成基因(PGC-1α/TFAM)及底物依賴性(Mitoplate S-1)。
關鍵靶點驗證:
藥理學(UK5099/羅格列酮抑制MPC)和遺傳學(siRNA沉默MPC1/MPC2)干預,結合FK866處理。
呼吸鏈復合物活性及ATP合成酶功能檢測。
機制整合:
解析丙酮酸→琥珀酸代謝轉換在維持膜電位(Δψm)和能量供應中的作用。
測量數據及其意義
(1)耐藥表型與機制排除(圖1)
數據:RES細胞對FK866完全耐藥(EC??不可計算),但對化療藥物(5FU、順鉑)敏感;Verapamil/CsA處理不逆轉耐藥;ABCB1/ABCC1表達無差異。
意義:耐藥非經典藥泵介導,提示代謝適應主導。
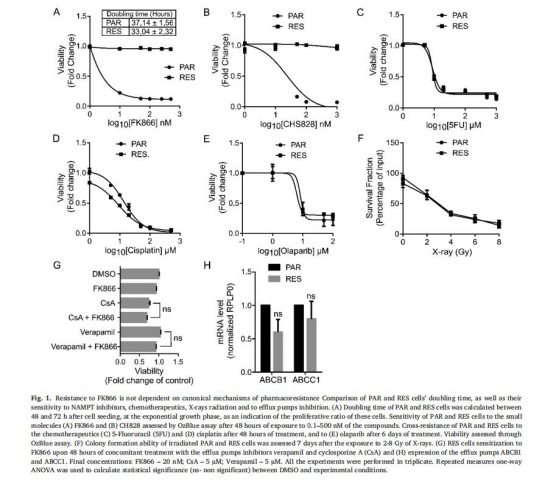 來源:圖1A-G。
來源:圖1A-G。
(2)NAD?合成通路無補償作用(圖2)
數據:RES細胞NAMPT/NAPRT下調、NMRK1上調,但NMRK1沉默不增敏FK866;NAD(H)/ATP水平不變。
意義:補償性NAD?合成非關鍵耐藥機制。
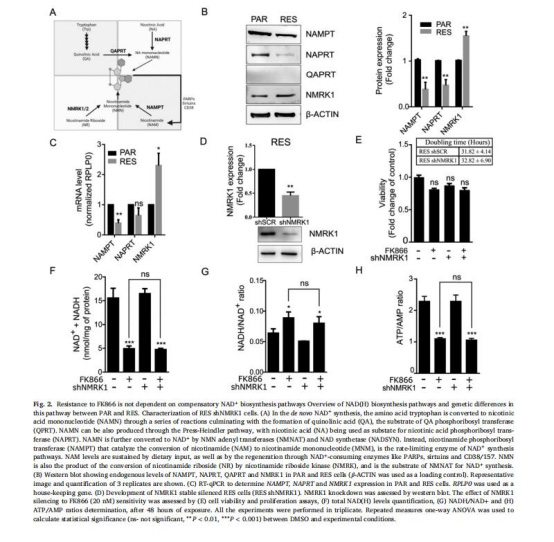
來源:圖2B-H。
(3)線粒體功能增強(圖3)
數據:
RES細胞備用呼吸容量(SRC) 顯著升高(Seahorse OCR)。
線粒體質量增加:mtDNA/nDNA比例↑、TOMM20蛋白↑、PGC-1α/TFAM轉錄上調、MitoTracker熒光強度↑2倍。
意義:線粒體生物合成與功能擴容是核心適應策略。

來源:圖3A-G。
(4)代謝底物依賴性轉變(圖4)
數據:Mitoplate分析顯示RES細胞對丙酮酸+低劑量蘋果酸氧化速率↑;UK5099(MPC抑制劑)降低RES細胞最大呼吸。
意義:耐藥細胞依賴丙酮酸作為碳源,MPC介導的丙酮酸輸入至關重要。
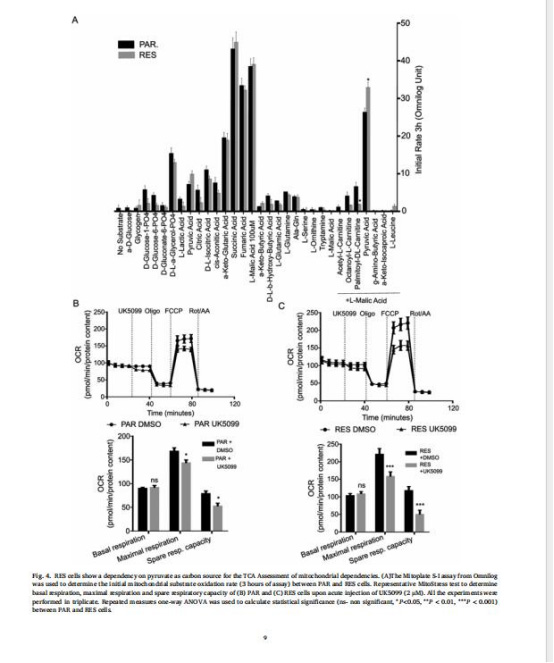
來源:圖4A-C。
(5)MPC2抑制誘導類耐藥表型(圖5)
數據:
UK5099/羅格列酮聯合FK866提升敏感細胞存活率。
MPC2沉默(非MPC1)部分挽救FK866導致的細胞死亡和ATP耗竭。
意義:MPC2功能抑制模擬耐藥表型,驗證其介導代謝適應的必要性。
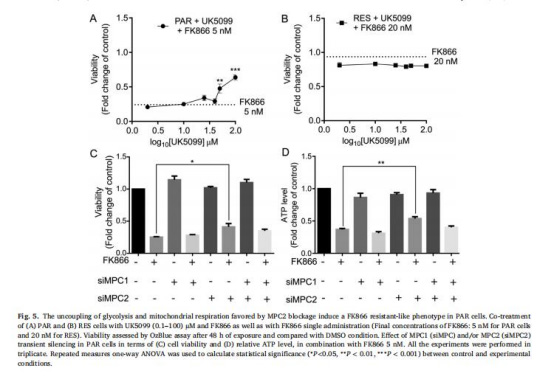
來源:圖5A-D。
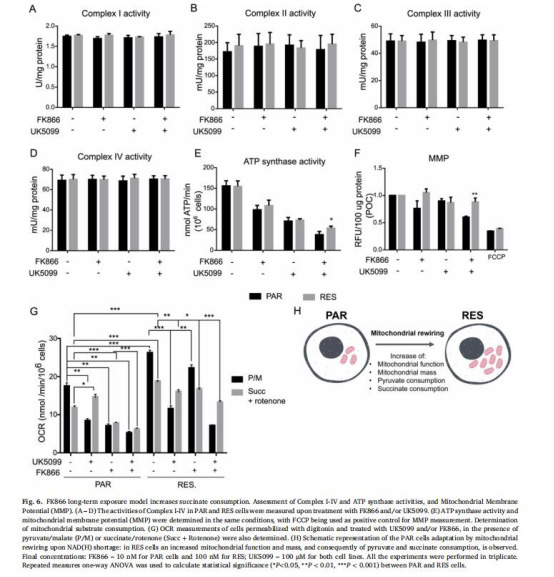
(6)琥珀酸依賴性與能量維持(圖6)
數據:
呼吸鏈復合物I-IV活性無差異,但ATP合成酶活性在FK866+UK5099處理下RES細胞更高。
丹麥Unisense微電極數據:RES細胞在丙酮酸剝奪(UK5099)下,琥珀酸氧化耗氧率(OCR) 顯著高于敏感細胞(圖6G)。
意義:耐藥細胞通過琥珀酸氧化維持膜電位(Δψm)和ATP合成,提供代謝靈活性。
來源:圖6A-G。
研究結論
線粒體代謝重組是耐藥核心:FK866持續暴露誘導線粒體生物合成(質量↑)和功能擴容(SRC↑),增強OXPHOS能力。
底物靈活性驅動適應:耐藥細胞從丙酮酸依賴轉向琥珀酸高效利用(Unisense數據證實),維持能量穩態。
MPC2是關鍵調控點:抑制MPC2(非MPC1)誘導敏感細胞產生類耐藥表型,靶向MPC2或可逆轉化療耐藥。
丹麥Unisense微電極數據的詳細解讀
測量方法與結果(圖6G)
技術原理:Unisense微電極通過實時耗氧率(OCR) 檢測,評估線粒體底物氧化效率。細胞經皂苷透化后,分別提供:
丙酮酸+蘋果酸(激活復合物I通路)。
琥珀酸+魚藤酮(激活復合物II通路,抑制復合物I)。
關鍵發現:
耐藥細胞琥珀酸OCR更高:RES細胞在琥珀酸底物下OCR顯著高于PAR細胞(*p<0.01),表明其增強琥珀酸氧化能力。
代謝靈活性:UK5099抑制丙酮酸輸入后,RES細胞仍通過琥珀酸維持高OCR;FK866處理不影響其琥珀酸利用效率。
敏感細胞的補償失敗:PAR細胞在丙酮酸剝奪后琥珀酸OCR無代償性提升,且FK866進一步抑制呼吸功能。
研究意義
揭示代謝可塑性:Unisense數據首次直接證明耐藥細胞通過琥珀酸氧化繞過丙酮酸短缺,維持線粒體呼吸,解釋其能量供應韌性。
提供耐藥新靶點:琥珀酸代謝途徑(如復合物II)或可成為克服NAMPT抑制劑耐藥的新干預方向。
技術優勢:Unisense微電極提供實時動態呼吸功能數據,彌補Seahorse靜態分析的不足,精準捕捉底物轉換的代謝適應性。
總結:本研究闡明TNBC細胞通過線粒體重編程(生物合成增強、底物靈活性提升)抵抗NAD(H)短缺導致的FK866毒性。丹麥Unisense微電極數據為核心機制(琥珀酸依賴)提供直接證據,為靶向線粒體代謝逆轉耐藥提供理論依據。