熱線:021-66110810,66110819
手機(jī):13564362870
熱線:021-66110810,66110819
手機(jī):13564362870
Resilience of recruiting seagrass (Thalassia testudinum) to porewater H?S in Florida Bay
佛羅里達(dá)灣海草對(duì)孔隙水 H 2 S 的恢復(fù)能力
來(lái)源:Aquatic Botany 187 (2023) 103650
1. 摘要核心內(nèi)容
研究對(duì)象:熱帶海草 Thalassia testudinum(海龜草)在佛羅里達(dá)灣高硫化氫(H?S)沉積物中的幼苗定植能力。
核心發(fā)現(xiàn):
幼苗抗H?S機(jī)制:幼苗通過(guò)(1)白天高效光合產(chǎn)氧、(2)夜間水柱擴(kuò)散供氧(潮汐輔助)、(3)根系發(fā)育延遲限制微生物產(chǎn)H?S,抵抗沉積物H?S脅迫。
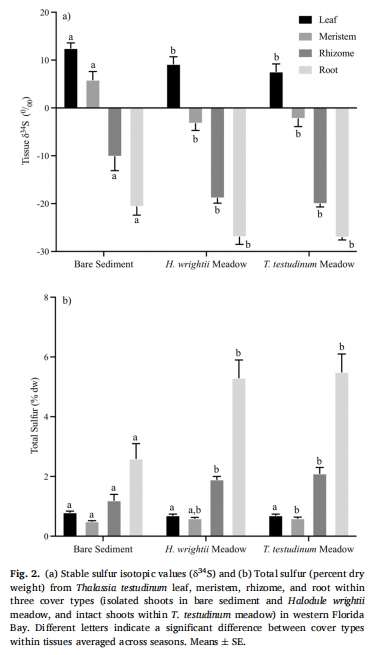
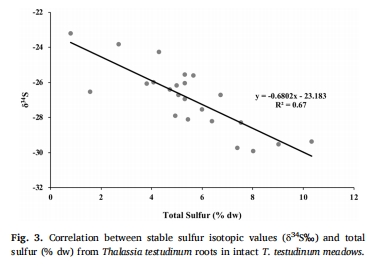
組織硫同位素(δ3?S):裸沙定植幼苗組織δ3?S值顯著高于成熟草甸(圖2a),表明其H?S暴露水平更低。
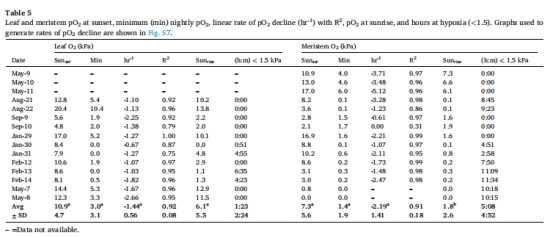
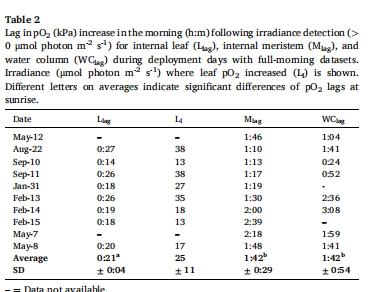
內(nèi)部氣體動(dòng)態(tài):Unisense微電極數(shù)據(jù)顯示,幼苗分生組織夜間缺氧時(shí)間長(zhǎng)于葉片(5:08 h vs 1:23 h),但未檢測(cè)到H?S積累(表5)。
結(jié)論:T. testudinum 幼苗可通過(guò)生理與發(fā)育策略在高H?S環(huán)境中成功定植,但草甸完全恢復(fù)需十年以上。
2. 研究目的
探究沉積物H?S是否限制 T. testudinum 幼苗向裸沙區(qū)域定植。
評(píng)估幼苗組織H?S暴露程度及內(nèi)部O?/H?S動(dòng)態(tài)。
揭示幼苗在極端環(huán)境(高H?S、缺氧)中的生存策略。
3. 研究思路
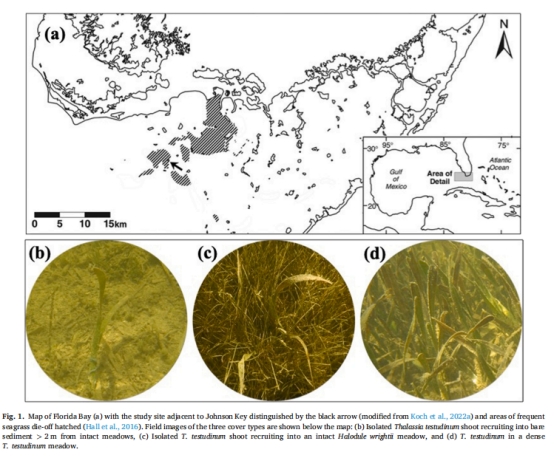
場(chǎng)地選擇:佛羅里達(dá)灣Johnson Key(圖1a),該區(qū)域因歷史性海草死亡事件形成大片裸沙區(qū)。
樣本設(shè)計(jì):對(duì)比三種生境:
裸沙定植幼苗(圖1b)
Halodule wrightii 草甸中的幼苗(圖1c)
成熟 T. testudinum 草甸(圖1d)
方法組合:
穩(wěn)定硫同位素(δ3?S):分析葉、分生組織、根莖、根組織硫源(圖2)。
Unisense微電極:實(shí)時(shí)監(jiān)測(cè)葉片/分生組織O?與H?S動(dòng)態(tài)(圖5)。
環(huán)境參數(shù):沉積物孔隙水H?S、水柱O?、光照、溫度等。
4. 測(cè)量數(shù)據(jù)及其意義
關(guān)鍵數(shù)據(jù)與來(lái)源
數(shù)據(jù)類(lèi)型 來(lái)源圖表 研究意義
組織δ3?S與總硫含量 圖2a-b 裸沙幼苗δ3?S更高(葉:+25‰,根莖:+50%),表明其H?S暴露低于草甸植株,根系發(fā)育延遲減少硫侵入。
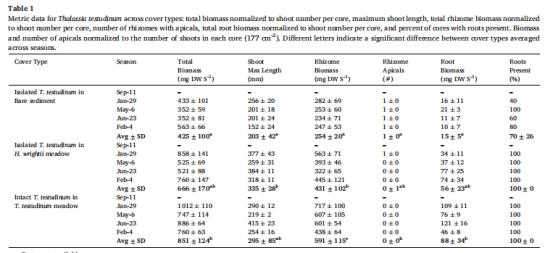
根系發(fā)育 表1 裸沙幼苗70%無(wú)根,根生物量比草甸低6倍,限制產(chǎn)H?S微生物群落。
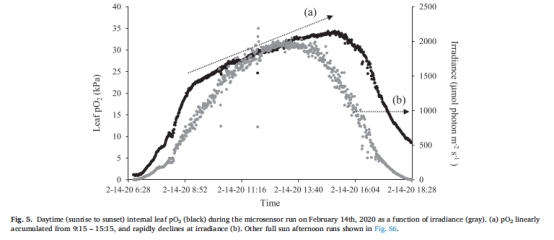
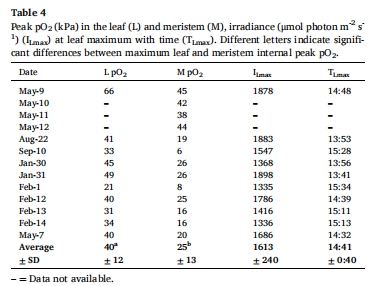
葉片/分生組織O?動(dòng)態(tài) 圖5、表5 幼苗葉片日間O?峰壓達(dá)40 kPa(海水飽和值2倍),分生組織夜間缺氧時(shí)間長(zhǎng)(5.1 h),但無(wú)H?S積累。
O?變化速率 表5 幼苗葉片O?上升速率(7 kPa/h)與草甸相當(dāng),但分生組織O?夜間下降更快(-2.19 kPa/h)。
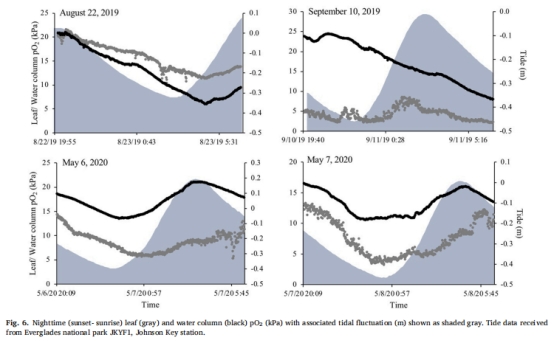
水柱O?與潮汐關(guān)聯(lián) 圖6 夜間漲潮提升水柱O?(>7 kPa),通過(guò)葉片擴(kuò)散補(bǔ)充內(nèi)部O?,緩解分生組織缺氧。
5. 核心結(jié)論
幼苗抗H?S機(jī)制:
日間氧化:高效光合在分生組織維持高O?(25 kPa),抑制H?S入侵。
夜間潮汐輔助:水柱O?通過(guò)葉片擴(kuò)散(圖6),抵消夜間缺氧風(fēng)險(xiǎn)。
根系策略:延遲根系發(fā)育減少沉積物微生物產(chǎn)H?S(表1)。
恢復(fù)潛力:幼苗可在高H?S裸沙定植,但草甸完全恢復(fù)需>10年。
管理意義:保護(hù)潮汐動(dòng)態(tài)與水質(zhì)是支持海草恢復(fù)的關(guān)鍵。
6. Unisense電極數(shù)據(jù)的深度解讀
測(cè)量方法
技術(shù):Unisense O?/H?S微電極插入葉片和分生組織(175 μm深度),同步記錄光照、溫度、水柱O?。
校準(zhǔn):
O?電極:100%飽和海水 vs 無(wú)氧抗壞血酸溶液兩點(diǎn)校準(zhǔn)。
H?S電極:0–1000 μM Na?S梯度校準(zhǔn)(pH=7.5確保H?S形態(tài))。
創(chuàng)新點(diǎn):首次在野外實(shí)時(shí)量化幼苗內(nèi)部O?/H?S動(dòng)態(tài),揭示潮汐對(duì)內(nèi)部氧化的調(diào)控作用。
關(guān)鍵結(jié)果(圖5, 表5)
日間O?動(dòng)態(tài):
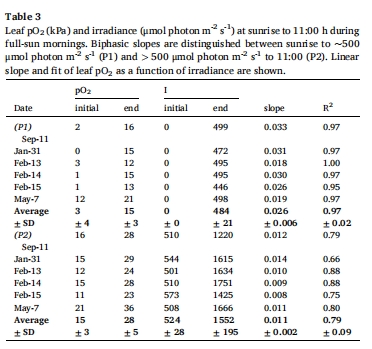
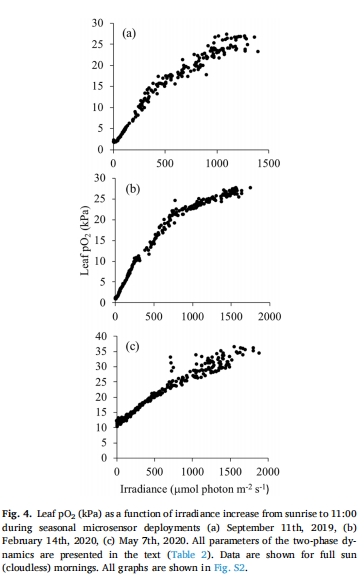
葉片O?在光照后20分鐘內(nèi)快速上升(7 kPa/h),峰壓達(dá)40 kPa(圖5)。
分生組織O?上升滯后1.7小時(shí),峰壓為葉片的60%(25 kPa),但仍足以抑制H?S積累。
夜間風(fēng)險(xiǎn):
分生組織夜間缺氧(<1.5 kPa)持續(xù)5.1小時(shí)(表5),但未檢測(cè)到H?S。
葉片O?通過(guò)水柱擴(kuò)散維持(3–10 kPa),漲潮時(shí)提升50%(圖6)。
H?S檢測(cè):所有監(jiān)測(cè)中分生組織H?S=0,印證δ3?S顯示的低暴露。
研究意義
機(jī)制驗(yàn)證:直接證明幼苗通過(guò)“O?緩沖區(qū)”抵御H?S,挑戰(zhàn)傳統(tǒng)“高H?S限制恢復(fù)”假說(shuō)。
潮汐作用:揭示水柱O?夜間通過(guò)葉片擴(kuò)散是內(nèi)部氧化的關(guān)鍵途徑(圖6)。
恢復(fù)策略:幼苗根系延遲發(fā)育是適應(yīng)性策略(非缺陷),為海草修復(fù)提供新視角——優(yōu)先保護(hù)幼苗定植期環(huán)境穩(wěn)定。
技術(shù)優(yōu)勢(shì):Unisense的高時(shí)空分辨率捕捉O?脈沖變化(如晨間快速上升),這是破壞性采樣無(wú)法實(shí)現(xiàn)的。
總結(jié)
本研究通過(guò)整合Unisense微電極、δ3?S同位素和發(fā)育生物學(xué)方法,揭示 T. testudinum 幼苗在高H?S環(huán)境中的三重抗性機(jī)制:日間光合產(chǎn)氧、夜間潮汐供氧、根系發(fā)育延遲。Unisense數(shù)據(jù)為核心結(jié)論提供直接證據(jù)——盡管分生組織夜間長(zhǎng)期缺氧,但高效氧化和潮汐輔助阻止H?S積累。該研究為海草生態(tài)修復(fù)提供了關(guān)鍵生理依據(jù):維持水柱O?和自然潮汐動(dòng)態(tài)比直接降低沉積物H?S更為重要。