熱線:021-66110810,66110819
手機:13564362870
熱線:021-66110810,66110819
手機:13564362870
結(jié)果
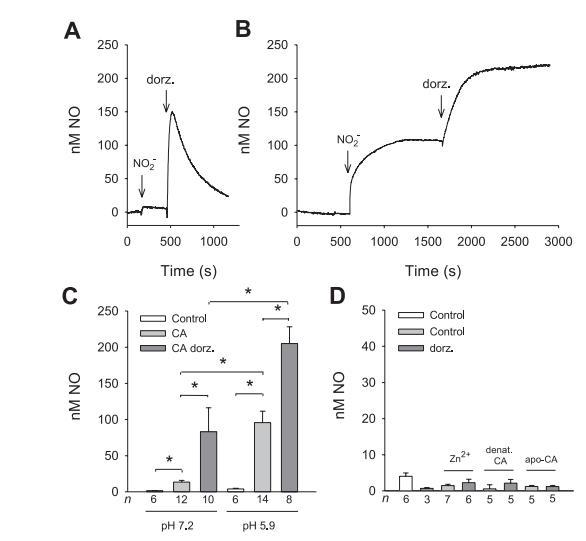
圖1多佐胺(Dorzolamide)和低pH會增加碳酸酐酶(CA)依賴亞硝酸鹽產(chǎn)生的一氧化氮(NO)。
使用微型一氧化氮電極來檢測在沒有和有純化CA(100μM)的情況下,在通氣磷酸鹽緩沖溶液中加入KNO2(100μM)后的NO產(chǎn)生情況(圖1A和B)。我們選擇的pH值分別為7.2和5.9,因為它們分別定義了缺血5-10分鐘后從正常細胞內(nèi)pH到組織pH的范圍。
正如預(yù)期的那樣,由于CA的反應(yīng)速率極低,在未催化的反應(yīng)中幾乎不會產(chǎn)生NO,而在有CA存在的情況下,加入亞硝酸鹽后很容易觀察到明顯更高的NO生成,尤其是在低pH值時(圖1)。為了抑制CA在NO生成反應(yīng)中的活性,我們添加了多佐胺(250μM),這是一種由CA催化的二氧化碳水合反應(yīng)的特異性抑制劑。之所以選擇這種磺酰胺,是因為它的溶解度高,而且不會干擾一氧化氮微電極,而另一種CA的特異性抑制劑乙酰唑胺的水溶性較低。
令人驚訝的是,加入多佐胺后,CA催化的NO生成量顯著增加(pH值分別為7.2和5.9時的6倍和2倍,圖1)。NO信號反映了NO生成和NO消耗之間的平衡,在pH值為7.2時,NO信號是短暫的,而在pH值為5.9時,NO信號延長(圖1A和B),最大NO信號被用于定量分析(圖1C)。在不同pH值下獲得的不同示蹤曲線表明,在反應(yīng)過程中,中間產(chǎn)物或產(chǎn)物種類在高pH值下被消耗(圖1A),或亞硝酸鹽在低pH值下再生(圖1B),但其機制仍有待研究。熱變性或Apo-CA酶(100μM)、Zn2+(以ZnCl2的形式添加,100μM)或單獨的多佐胺(250μM)不會導(dǎo)致100μM亞硝酸鹽產(chǎn)生的NO顯著增加(圖1D)。總之,這些數(shù)據(jù)表明,CA能夠與亞硝酸鹽反應(yīng)生成NO,而且在低pH值和多佐胺存在的情況下,產(chǎn)生的NO水平更高。
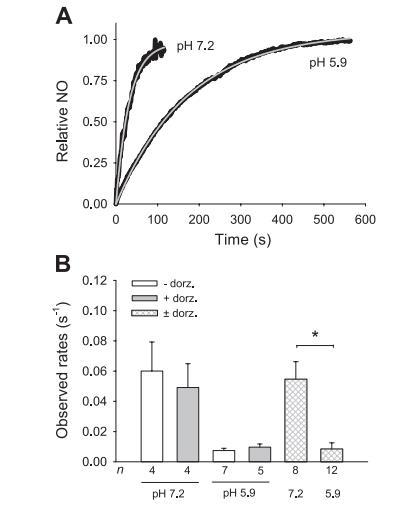
圖2亞硝酸鹽與CA的反應(yīng)動力學(xué)。
為了確定亞硝酸鹽在CA催化反應(yīng)中產(chǎn)生NO的速率,我們分析了在沒有多佐胺和有多佐胺的情況下,向CA溶液中加入最終濃度為100μM的亞硝酸鹽(圖2A)后,使用一氧化氮電極收集的動力學(xué)軌跡。
只分析了最慢的反應(yīng)軌跡,這表明注入的亞硝酸鹽已在反應(yīng)室中完全混合。由于反應(yīng)是在二階條件下研究的,即在酶和底物濃度相等(100μM)的條件下,NO的動力學(xué)跡線可以用雙曲線函數(shù)很好地擬合(r2=0.981±0.024,n=20)。單指數(shù)函數(shù)的擬合效果同樣良好(r2=0.975±0.084,n=20)(圖2A),表明游離底物亞硝酸鹽的濃度超過了與酶結(jié)合的亞硝酸鹽的濃度,反應(yīng)是在假一階條件下進行的。這一發(fā)現(xiàn)與報道的亞硝酸鹽對CA的低親和力相一致。
觀察到的速率明顯受到pH值的影響,但不受多佐胺的影響(圖2B),根據(jù)單指數(shù)擬合得到的值計算得出的表觀速率常數(shù)(平均±SE)為:pH值為7.2時547±116/M/s,pH值為5.9時84±42/M/s(圖2B)。由于pH值為7.2時反應(yīng)的半衰期(t1/2,~13秒)與電極的反應(yīng)時間(~10秒)相當(dāng),因此該pH值下的反應(yīng)速率快于~547/M/s。
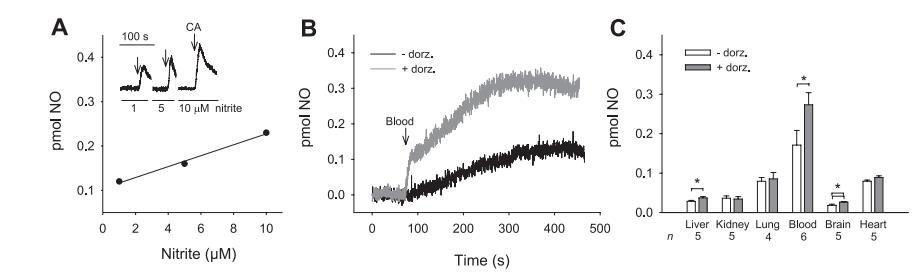
圖3純化CA和大鼠組織在37℃、采樣率為4s-1的條件下由亞硝酸鹽產(chǎn)生NO的化學(xué)發(fā)光分析。
在動物組織中發(fā)現(xiàn)的酶和亞硝酸鹽濃度較低時,采用高靈敏度的NO化學(xué)發(fā)光方法測量NO的產(chǎn)生,該方法可在用純凈N2吹掃反應(yīng)容器時檢測氣相中的NO。與使用檢測溶液中NO的一氧化氮電極進行的實驗不同,檢測氣相中NO的化學(xué)發(fā)光法還能監(jiān)測N2O3的產(chǎn)生。
在本文研究的反應(yīng)過程中會形成N2O3(見公式2),當(dāng)在氣相中凈化時,N2O3會完全均解,生成NO。如圖3A所示,在含有pH值為7.2的磷酸鹽緩沖液中的亞硝酸鹽(1、5和10μM)的NO分析儀的凈化容器中注入10μM的純化CA時,觀察到了依賴亞硝酸鹽的NO生成。這證實了圖1中的數(shù)據(jù),并進一步表明該反應(yīng)也發(fā)生在厭氧條件下。然后,我們在pH值為5.9、含有100μM亞硝酸鹽的凈化容器中注入大鼠組織勻漿后檢測了NO的產(chǎn)生。在這些條件下,亞硝酸鹽轉(zhuǎn)化為NO的許多途徑都有助于檢測到NO。
為了區(qū)分由組織特異性CA從亞硝酸鹽產(chǎn)生的NO和由其他途徑產(chǎn)生的NO,我們在沒有多佐胺和有多佐胺的情況下進行了化學(xué)發(fā)光分析,以檢測多佐胺是否對各組織產(chǎn)生的NO有顯著影響,多佐胺能與CA發(fā)生特異性反應(yīng),從而增強亞硝酸鹽產(chǎn)生NO的能力,如圖1所示。在所有組織中都能檢測到亞硝酸鹽產(chǎn)生的NO,這與之前的研究結(jié)果一致,血液、肝臟和大腦在存在多佐胺的情況下產(chǎn)生的NO顯著增加(圖3B和C)。特別是,在含有和不含多佐胺的血液中觀察到的高NO信號(圖3,B和C)表明,亞硝酸鹽依賴性NO的生成超過了血紅蛋白的NO清除能力,這與之前的觀察結(jié)果一致。由于單獨使用多佐胺對NO的生成沒有影響(圖1),多佐胺的這種增強效應(yīng)表明,組織中(至少在血液、肝臟和大腦中)存在能夠催化亞硝酸鹽轉(zhuǎn)化為NO的功能性CA,盡管CA同工型和其他酶在生理條件下不同組織將亞硝酸鹽轉(zhuǎn)化為NO的能力中的相對作用仍有待解決。
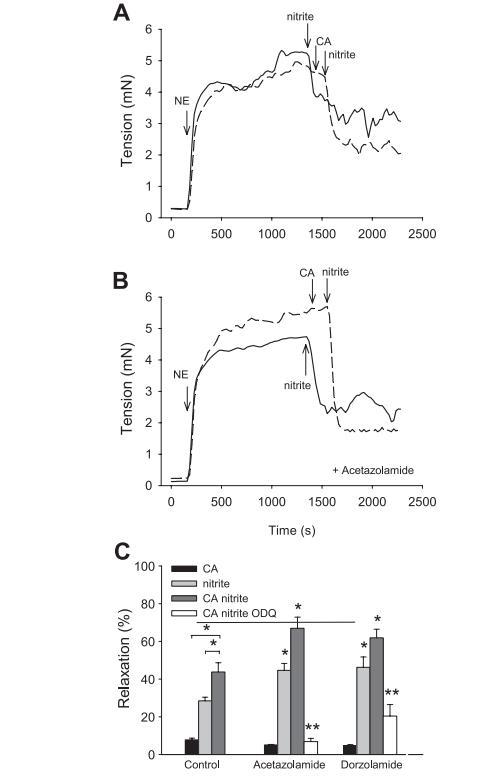
圖4亞硝酸鹽與CA反應(yīng)產(chǎn)生的NO的血管活性以及乙酰唑胺、多佐胺和1H-[1,2,4]oxadiazolo[4,3a]quinoxalin-1-one(ODQ)的影響。
為了驗證在低亞硝酸鹽(~10μM)和高HCO3-(~25mM,5%CO2,pH7.4)的生理條件下,CA產(chǎn)生的NO是否具有血管活性,我們按照之前的描述,在1%O2和5%CO2條件下進行了大鼠主動脈環(huán)生物測定(圖4A和B)。在這些條件下,亞硝酸鹽的濃度為10μM,接近主動脈血管中的濃度,能夠誘導(dǎo)50%的血管擴張。在不添加亞硝酸鹽的情況下,添加外源CA(10μM)對血管舒張僅有輕微影響,而添加CA和亞硝酸鹽所引起的血管舒張明顯大于單獨使用CA或亞硝酸鹽所測得的結(jié)果(圖4C)。在乙酰唑胺和多佐胺存在的情況下,大鼠胸主動脈段的亞硝酸鹽依賴性血管舒張明顯增加(圖4C)。相反,在有ODQ和乙酰唑胺或多佐胺存在的情況下,加入CA和亞硝酸鹽誘導(dǎo)的血管舒張明顯減弱,這表明觀察到的舒張效應(yīng)主要依賴于cGMP。這些數(shù)據(jù)表明,CA與生理水平的亞硝酸鹽反應(yīng),通過激活可溶性鳥苷酸環(huán)化酶產(chǎn)生具有血管活性的NO。
 相關(guān)新聞
相關(guān)新聞